Tadalafil appartiene alla classe degli inibitori selettivi della fosfodiesterasi di tipo 5, con un profilo farmacocinetico caratterizzato da un’emivita terminale di circa diciotto ore. Dopo somministrazione orale viene assorbito rapidamente e raggiunge concentrazioni plasmatiche massime in due ore. La biotrasformazione avviene principalmente tramite CYP3A4 con formazione di metaboliti inattivi, escreti in prevalenza con le feci. L’elevato legame alle proteine plasmatiche (>90%) assicura una distribuzione stabile. Nei confronti delle altre molecole della stessa classe, cialis compresse italia è noto per la durata prolungata dell’attività farmacologica.
Ejn_6201 2466.2472
European Journal of Neuroscience, Vol. 27, pp. 2466–2472, 2008
Augmented brain 5-HT crosses the blood–brain barrierthrough the 5-HT transporter in rat
Yasushi Nakatani, Ikuko Sato-Suzuki, Naohisa Tsujino, Akane Nakasato, Yoshinari Seki, Masaki Fumotoand Hideho AritaDepartment of Physiology, Toho University School of Medicine, 5-21-16, Omori-nishi, Ota-ku, Tokyo 143–8540, Japan
Keywords: blood–brain barrier, microdialysis, serotonin, SSRI
The present study re-evaluated an existing notion that serotonin (5-hydroxytryptamine; 5-HT) could not cross the brain to thecirculating blood via the blood–brain barrier (BBB). To elevate brain 5-HT alone, 5-hydroxytryptophan (5-HTP; 30–75 mg ⁄ kg) wasadministrated intravenously to anaesthetized rats that had undergone gastrointestinal and kidney resections along with liverinactivation (organs contributing to increasing blood 5-HT after 5-HTP administration). A microdialysis method and HPLC systemwere used to determine the brain 5-HT levels in samples collected from the frontal cortex. Blood 5-HT levels were determined fromwhole blood, not platelet-poor plasma, collected from the central vein. We found that blood 5-HT levels showed a significantaugmentation whenever brain 5-HT levels were significantly elevated after the administration of 5-HTP in those rats with theabdominal surgical procedures. This elevation was abolished after pretreatment with a selective serotonin reuptake inhibitor(fluoxetine; 10 mg ⁄ kg i.v.), although brain 5-HT levels remained augmented. These results indicate that augmented brain 5-HT cancross the BBB through the 5-HT transporter from the brain to the circulating blood.
It was previously believed that serotonin (5-hydroxytryptamine;
brain ECF 5-HT levels in this study. As 5-HTP decarboxylase, which
5-HT) could not cross from the brain to the periphery via the
metabolizes 5-HTP to 5-HT, is found not only in the brain but also
blood–brain barrier (BBB). However, recent in vitro studies (Brust
in the gut, liver and kidneys (West, 1958), a rise in blood 5-HT
et al., 2000; Wakayama et al., 2002) have revealed the presence of
levels after 5-HTP administration may not be due to the brain alone
5-HT transporter mRNA in vascular endothelial cells, indicating
but also to these other organs. Therefore, to exclude this possibility
that the BBB may act as an efflux transport system for 5-HT. Based
we surgically removed the potentially complicating organs. We
on this information, we conducted the present study to re-evaluate
administered 5-HTP to those rats whose gastrointestinal tracts and
the above concept. In other words, we examined the possibility that
kidneys had been completely resected and whose livers had been
augmented brain 5-HT may cross the BBB through the 5-HT
To confirm the role of the BBB 5-HT transporter in the transport of
5-HT neurons, which are located in the raphe nuclei of the
5-HT from the brain to the periphery, we also examined the effect of
brainstem, are known to project to broad areas of the whole brain. In
selective serotonin reuptake inhibitors (SSRIs) in those rats. Brain
addition, 5-HT neuronal activity exhibits a state-dependent change
ECF 5-HT and blood 5-HT levels were measured after 5-HTP
(Jacobs & Azmitia, 1992). These neurons regularly fire during waking,
administration in rats with and without SSRI pretreatment.
although the firing rate is irregular or silenced during sleep. Therefore,
However, it has been established that blood 5-HT is mainly
it can be anticipated that there is a steady synaptic release of 5-HT that
distributed in platelets and to a lesser degree in the plasma (Artigas
occurs over a broad area of the brain during waking. In fact, rat brain
et al., 1985; Ortiz et al., 1988). If brain ECF 5-HT can be transported
5-HT levels revealed state-dependent alterations, i.e. high levels
via the BBB, it would be expected that the augmented 5-HT in the
during waking and low levels during sleep (Penalva et al., 2003). In
plasma would move quickly into the platelets. Therefore, in this study
the present study, we evaluated the change in extracellular fluid (ECF)
we evaluated changes in whole blood, not in platelet-poor plasma, to
5-HT levels in the brain using a microdialysis method and a high-
evaluate the BBB efflux transport system for 5-HT from the brain to
performance liquid chromatography (HPLC) system.
It has been well established that brain 5-HT is elevated by
administration of 5-hydroxytryptophan (5-HTP), i.e. the 5-HTprecursor (Okada et al., 1972; Lo¨scher et al., 1984; Gartside
et al., 1992). We injected 5-HTP intravenously to rats to elevate
All procedures involving animals were carried out in accordance
Correspondence: Dr Hideho Arita, as above.
with the European Communities Council Directive of 24 November
1986 (86 ⁄ 609 ⁄ EEC) and were approved by the Animal Experimen-
Received 26 November 2007, revised 28 February 2008, accepted 11 March 2008
tation Ethics Committee of the Toho University School of Medicine.
ª The Authors (2008). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing Ltd
Experiments were performed on 25 male Wistar rats weighing 310–
at a flow rate of 0.50 mL ⁄ min. The column temperature was
375 (327.6 ± 4.2) g. The rats were adapted to the standard 12 : 12 h
light : dark conditions (lights on at 08.00 h) for > 1 week before the
After the stabilization period, three consecutive measurements of
experiments. Experiments were then performed during the light
5-HT, which were carried out every 10 min, were done to confirm
cycle. The rats were anaesthetized with 50 mg ⁄ kg pentobarbital
that the microdialysis sampling from the FC was exhibiting a
sodium intraperitoneally. The depth of anaesthesia was continuously
steady-state baseline. Thereafter, the three different experimental
controlled by maintaining the absence of nociceptive flexion and
procedures using 5-HTP and ⁄ or SSRI were carried out (described
corneal reflexes. Supplementary doses (20% of the original dose)
were given intravenously when necessary. The anaesthesia wasmaintained until the end of the experiment, when the animal waskilled with an overdose of anaesthetic. All efforts were made to
minimize the number of animals used.
Blood (0.5 mL) was obtained in a plastic tube and then 0.5 mL of
Catheters were placed into the central vein near the right atrium for
saline solution was injected back into the animal in order to maintain a
collection of blood samples and for drug injection. To monitor vital
stable and constant total blood amount. As per the method of Kremer
signs, we measured the arterial blood pressure by placing a catheter
et al. (1990), 0.5 mL of blood was suspended in 2.2 mL of water.
into the femoral artery. Heart rate was calculated from the blood
Then, 300 lL of the internal standard and 10 lL of a 10% (weight per
pressure pulse with a tachometer (AT-601G; Nihon Kohden). The
volume) solution of ascorbic acid in water was added to the suspended
animals were fixed in a prone position in a stereotaxic frame. Rectal
blood sample. The sample was then frozen at )20 °C and stored until
temperature was maintained at 37 °C with a heating lamp.
Rats used in this study had undergone total resection of the
5-HT analyses of the blood samples were conducted within
gastrointestinal tract and kidneys, as well as liver inactivation. In
1 week after the experiment. Blood samples were thawed, and then
these rats, the operative area was shaved and the abdomen opened by
167 lL of methanol was added to 1 mL of blood sample in order
a long midline incision with aseptic precautions. Ligatures were
to remove the proteins. Blood samples were centrifuged at 4670 g
placed round the celiac and the superior mesenteric arteries as near
for 10 min at 4 °C. Five hundred microlitres of the supernatant of
to the aorta as possible. To inactive the liver, the portal vein was
the blood sample was suspended in 4.5 mL of the mobile buffer.
ligated along with the bile duct. The whole intestine was freed from
Twenty microlitres of the blood sample was then injected into the
its ligaments and all vascular attachments were removed. Total
HPLC system. Although we used the same HPLC system to
resection of the gastroduodenum and the small and large intestines
determine blood 5-HT levels, we applied the following different
was performed as quickly and carefully as possible to minimize
method for blood 5-HT analysis from that for FC 5-HT analysis.
bleeding. After ligation, the kidneys were also removed. The rectum
with its vessels was clamped, ligated distally to the clamp, and then
CA-5ODS; 2.1 mm diameter · 150 mm; Eicom). The mobile phase
cut between the clamp and the point of ligation. Abdominal muscles
consisted of a 0.1-m phosphate acid buffer containing 50 mg ⁄ L
and skin were then sutured. After the operation, we confirmed that
EDTAÆ2Na and 300 mg ⁄ L sodium 1-octanesulphonate (Nacalai
the animal was breathing spontaneously and that the blood pressure
Tesque, Japan) as the ion-pair and 20% methanol at pH 6.0. The
was maintained within the normal range (mean arterial pressure
flow rate was set at 0.22 mL ⁄ min and the column temperature was
121.45 ± 4.11 mmHg; n ¼ 20). The rats were fixed in a prone
Microdialysis procedures and 5-HT measurements
A parietal craniotomy was performed, and the dura was opened toadvance the microdialysis probe (0.22 mm diameter; 2 mm exposed
The first experiment was performed on five intact rats that did not
membrane; A-I-4–02; Eicom, Japan). Using the atlas of the rat brain
undergo the abdominal operation. After the stabilization period of
(Paxinos & Watson, 1986) as a guide, a probe was slowly and gently
1–2 h, we determined the steady-state level of brain 5-HT by
inserted into the left frontal cortex (FC), an area 0.5 mm lateral to the
measuring three consecutive microdialysis perfusate samples from
midline, 3.2 mm anterior to the bregma and 2.5 mm vertically below
the FC at 10-min intervals. Thereafter, 1 mL of 5-HTP (a dose of
the dura. The probe was fixed with dental cement, connected to a
75 mg ⁄ kg in saline solution) was administered through the venous
microinfusion pump (EP-50; Eicom) and then perfused with Ringer’s
Microdialysis sampling from the FC was carried out every 10 min
m; KCl, 4 mm; CaCl2, 1.9 mm) at a flow rate of
throughout the entire experiment. Blood sampling was performed four
A stabilization period of at least 1 h was allowed following probe
times as follows. The first blood sample was taken prior to the 5-HTP
implantation. After the stabilization period, microdialysis sampling
administration. Following the 5-HTP administration three further
from the FC was carried out every 10 min. The perfusate from the FC
blood samples were drawn at 1-h intervals. Note that same amount of
was injected into a HPLC system (DAM-300 system; Eicom) using an
saline solution was injected back into the animals in order to maintain
automatic injector (AS-10; Eicom) and immediately analysed for
a stable and constant total blood amount.
5-HT. A reverse-phase column (Eicompak PP-ODS, 4.6 mm diame-ter · 30 mm; Eicom) was used for 5-HT separation. The working
electrode was a graphite electrode set at a detector potential of
The second experiment was performed on 10 rats that underwent
+0.40 V against an Ag ⁄ AgCl2 reference electrode. The composition of
complete resection of their gastrointestinal tracts and kidneys, along
the mobile phase was 0.1 m phosphoric acid buffer at pH 6.0,
with liver inactivation. One millilitre of 5-HTP (a dose of 75 mg ⁄ kg in
containing 1% methanol, 2 mm sodium 1-decanesulphonate (as the
saline solution) was administered through a venous catheter in six rats,
ion-pair) and 0.13 mm ethylenediaminetetraacetic acid (EDTAÆ2Na)
with the same amount of a saline solution injected in four rats that
ª The Authors (2008). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 27, 2466–2472
served as the control animals. Following the injection of 5-HTP or
saline solution, samples were obtained from the FC every 10 min for
Experiment 1: effect of 5-HTP administration on the frontal
cortex (FC) and blood 5-HT levels in intact rats
Blood sampling was performed three times in the six rats given
5-HTP. The first blood sample was taken prior to the injection of
The first experiment was performed in intact rats that did not undergo
5-HTP (preinjection sample). The second blood sample was obtained
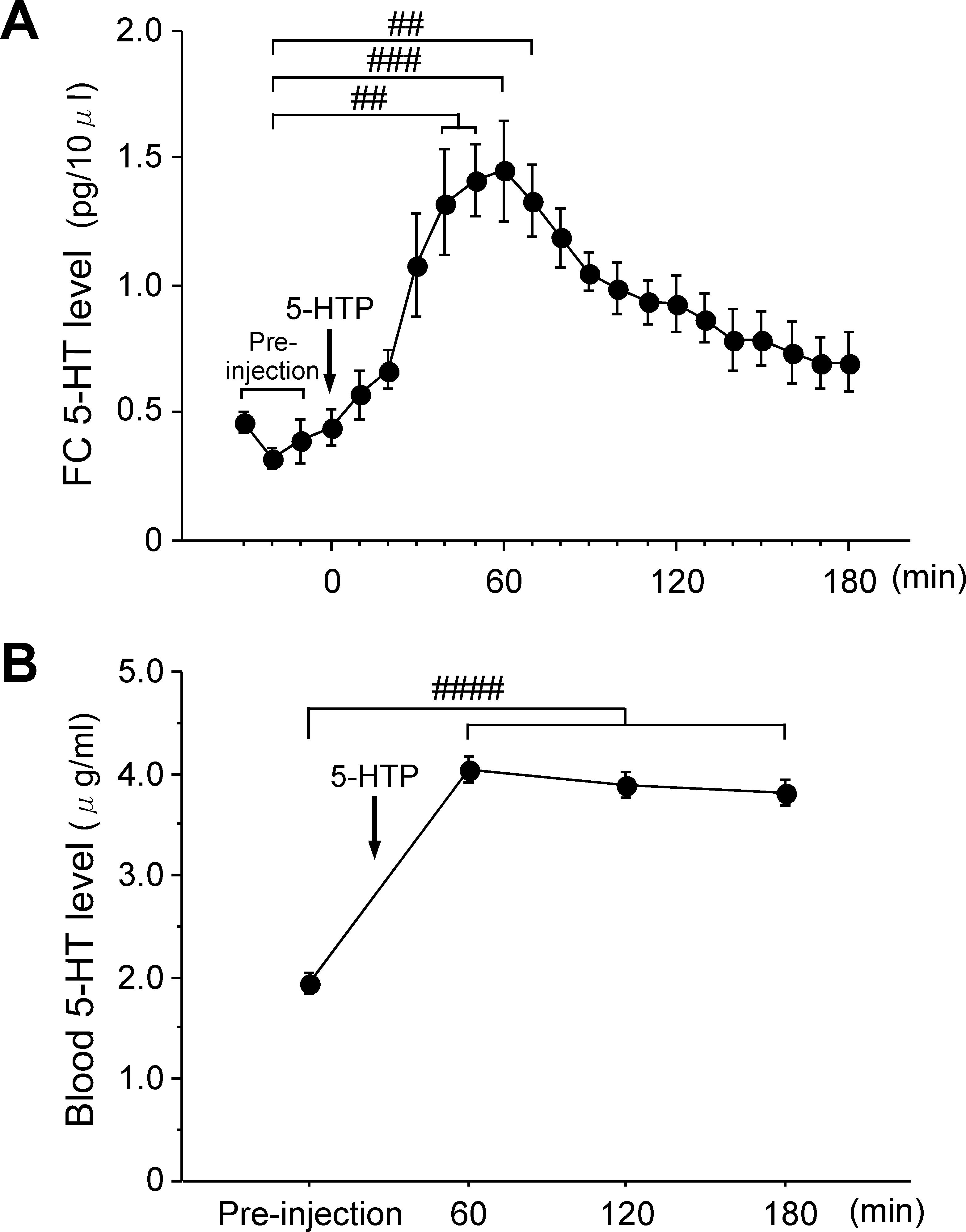
the abdominal operation. Figure 1A shows the statistical data for the
when the brain 5-HT level had increased to more than two times
time course of the changes in FC 5-HT levels before and after 5-HTP
higher than the preinjection level, which occurred at 50–70 min after
administration. The mean preinjection 5-HT level in FC was
5-HTP administration. The third blood sample was taken when the
0.39 ± 0.04 pg ⁄ 10 lL (n ¼ 5). When 5-HTP was administered
brain 5-HT level further increased, which occurred at 10–30 min after
intravenously, there was a gradual increase in FC 5-HT levels.
A one-way anova revealed significant changes for the time course of
In the control experiment, three consecutive blood samples were
the mean FC 5-HT level after 5-HTP administration (F4,19 ¼ 9.29,
drawn before the saline solution administration and then every 1 h
P < 0.0001). There was a significant post hoc difference after 5-HTP
after saline administration in the four rats that were not given 5-HTP.
administration between before and 40 min (P < 0.01), 50 min(P < 0.01), 60 min (P < 0.001) and 70 min (P < 0.01) after 5-HTP
administration. The maximum FC 5-HT level observed at 60 minafter the 5-HTP injection was 1.45 ± 0.44 pg ⁄ 10 lL (n ¼ 5), which
The third experiment was conducted to evaluate the effect of SSRI
was approximately three times higher than the mean preinjection
(fluoxetine hydrochloride; Sigma, USA). The experiment was per-
level. FC 5-HT levels gradually decreased thereafter. The mean FC
formed on 10 rats that underwent resection of their gastrointestinal
tracts and kidneys along with liver inactivation. Prior to SSRI
pretreatment, we determined the steady-state level of the brain 5-HTby measuring three consecutive microdialysis perfusate samples fromthe FC every 10 min (pretreatment period). Thereafter, 1 mL of SSRI(a dose of 10 mg ⁄ kg in saline solution) was administered to ratsthrough a venous catheter. One millilitre of 5-HTP (a dose of30 mg ⁄ kg in saline solution) was administered through the venouscatheter at 40 min after the SSRI pretreatment. The dose of 5-HTPgiven was reduced to approximately half of that given in Experi-ments 1 and 2. The reason for the reduction in 5-HTP loading was thatextremely high levels of brain 5-HT were noted in the firstmicrodialysis perfusate sample from the FC when we administeredthe same 5-HTP dose (75 mg ⁄ kg in saline solution) with SSRIpretreatment in the animals.
Microdialysis sampling from the FC continued every 10 min
throughout the experiment, with blood samples drawn three timesduring the study period. The first blood sample was obtained duringthe control period, i.e. prior to SSRI pretreatment. The second bloodsample was drawn 30 min after SSRI pretreatment, which was 10 minprior to the 5-HTP administration. The third blood sample was drawnat the time when we observed an apparent increase in the FC 5-HTlevel after the 5-HT administration, which was 30–90 min after 5-HTPadministration.
In the control experiment, instead of SSRI pretreatment animals
were given 1 mL of saline solution through the venous catheterafter the control period, which was described above. One millilitreof 5-HTP was given through the venous catheter at 40 min afterthe saline pretreatment. Microdialysis sampling from the FCcontinued every 10 min throughout the experiment. Blood sampleswere drawn three times during the control experiment. The firstsample was drawn during the control period, the second 30 minafter saline pretreatment and the third 0–90 min after 5-HTPadministration.
Fig. 1. The effects of 5-HTP administration on (A) the brain and (B) the blood
5-HT levels in intact rats. An arrow indicates the timing of the 5-HTPadministration. (A) Time course showing changes in the mean 5-HT level in
A one-way anova was used to analyse the data obtained in
the FC before and after 5-HTP administration (75 mg ⁄ kg, i.v.) in the intact rats.
Experiment 1 while a two-way anova for repeated measures was
The preinjection level of the FC 5-HT consists of the mean of three successive
used for data obtained in Experiments 2 and 3. Significant main
data points obtained at 10, 20 and 30 min before 5-HTP administration. (B) Time course showing changes in the mean blood 5-HT level before and
effects were further analysed with a Scheffe´ post hoc test. Effects were
after 5-HTP administration (75 mg ⁄ kg, i.v) in the intact rats. Data are
considered to be statistically significant when P-values were < 0.05.
expressed as the mean ± SE (n ¼ 5). ##P < 0.01, ###P < 0.001, ####P < 0.0001
All data are expressed as the mean ± SE.
ª The Authors (2008). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing Ltd
European Journal of Neuroscience, 27, 2466–2472
Figure 1B shows the time course for the changes in blood 5-HT
at 60 min after 5-HTP administration. On the other hand, rat 4
levels. The mean blood 5-HT level before 5-HTP administration was
expressed a slow and small increase in the FC 5-HT level after 5-HTP
1.95 ± 0.10 lg ⁄ mL. A one-way anova revealed significant changes
administration, with a FC 5-HT level of 14.1 pg ⁄ 10 lL at 60 min and
in the time course for the mean blood 5-HT level after 5-HTP
a peak FC 5-HT level of 105.8 pg ⁄ 10 lL at 120 min after 5-HTP
administration (F4,3 ¼ 111.5, P < 0.0001). There was a significant
administration. As the time courses for the changes in FC 5-HT levels
post hoc difference between before and 60 min (P < 0.0001),
differed among the six rats after 5-HTP administration, we obtained
120 min (P < 0.001) and 180 min (P < 0.001) after 5-HTP admin-
the first postinjection blood sample at a time when the FC 5-HT level
istration. The maximum blood 5-HT level observed at 60 min after the
was more than two times higher than the preinjection level. Thus, the
5-HTP injection was 4.40 ± 0.14 lg ⁄ mL, which was approximately
first postinjection blood samples (open symbol for each response
two times higher than the preinjection blood 5-HT level. The blood
curve) were drawn between 50 and 70 min after 5-HTP administration
5-HT levels continued to remain high until 180 min after 5-HTP
in this experiment. The second postinjection samples were taken when
administration, although FC 5-HT levels showed an apparent decrease
the FC 5-HT level further increased, which occurred at 10–30 min
after the first postinjection blood samples were drawn.
Figure 2B shows the changes in FC 5-HT levels before and after
saline administration in four rats that had total removal of their
Experiment 2: effect of 5-HTP administration on FC 5-HT levels
gastrointestinal tracts and kidneys, along with liver inactivation. The
and blood 5-HT levels in rats that had undergone the abdominal
0.53 ± 0.07 pg ⁄ 10 lL. Little or no change in the FC 5-HT levels
The second experiment was performed in 10 rats that had undergone
was observed throughout the experiment and up to 120 min after
total removal of their gastrointestinal tracts and kidneys, along with
saline administration. Thus for these rats, sequential blood samplings
liver inactivation. Figure 2 shows the individual time course for the
were obtained at 60 and 120 min after saline administration.
changes in the FC 5-HT levels before and after 5-HTP or saline
Figure 3 shows the statistical data for the time course changes in the
administration. The mean FC 5-HT level during the preinjection
blood 5-HT levels after 5-HTP and saline administrations. The mean
period was 0.89 ± 0.19 pg ⁄ 10 lL (n ¼ 6). After 5-HTP was admin-
blood 5-HT level before 5-HTP administration was 2.14 ± 0.11
istered intravenously, there was a gradual increase observed in the FC
lg ⁄ mL (n ¼ 6). There was a gradual increase in the mean blood 5-HT
5-HT levels, although there were marked differences for the time
level after 5-HTP administration, whereas there was little change
course and magnitude of the changes in the FC 5-HT level among the
noted for the mean blood 5-HT level after saline administration.
six rats examined (Fig. 2A). For example, rat 2 exhibited a relatively
A two-way anova revealed significant changes in the time course for
rapid and large increase in the FC 5-HT level after 5-HTP
the mean blood 5-HT level after 5-HTP administration. The interaction
administration, with a peak FC 5-HT level of 186.3 pg ⁄ 10 lL seen
effect on the time course of the mean blood 5-HT level was significantfor 5-HTP administration · saline administration (F1,2 ¼ 43.65,P < 0.001). In addition, individual anova revealed a significantchange in the time course for the mean blood 5-HT level after 5-HTPadministration (F3,2 ¼ 50.41, P < 0.001), which was not seen aftersaline administration (F3,2 ¼ 0.77, P ¼ 0.50). With regard to themean blood 5-HT level after 5-HTP administration, there was a
Fig. 2. The effects of (A) 5-HTP and (B) saline administration on FC 5-HTlevel in rats that had undergone total removal of their gastrointestinal tracts andkidneys, along with liver inactivation. (A) Time courses showing changes inindividual FC 5-HT levels before and after 5-HTP administration in six rats that
Fig. 3. Time courses showing changes in the mean blood 5-HT levels before
had undergone the abdominal operation. The arrow indicates the timing of
and after 5-HTP (s, n ¼ 6) and saline (d, n ¼ 4) administrations in rats that
5-HTP administration (75 mg ⁄ kg, i.v). (B) Time courses showing changes in
had undergone total removal of their gastrointestinal tracts and kidneys, along
individual FC 5-HT levels before and after saline administration (arrow) in four
with liver inactivation. The arrow indicates the time of the 5-HTP or saline
rats that had undergone the abdominal operation. Open symbols indicate the
administration. Data are expressed as the mean ± SE. ###P < 0.001 vs.
times when blood samples were drawn.
preinjection values. *P < 0.05, **P < 0.01, 5-HTP vs. Saline.
ª The Authors (2008). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 27, 2466–2472
significant post hoc difference observed between the preinjectionsample and the first (P < 0.001) and second (P < 0.001) postinjectionsamples. There was also a significant post hoc difference between5-HTP administration and saline administration for both the first(P < 0.05) and second (P < 0.01) postinjection samples.
Experiment 3: combined effects of SSRI and 5-HTPadministrations on FC 5-HT levels and blood 5-HT levelsin rats that had undergone the abdominal operation
The third experiment was also performed in 10 rats that had undergonetotal removal of their gastrointestinal tracts and kidneys, along withliver inactivation. Figure 4A shows individual time courses for thechanges in FC 5-HT levels before and after 5-HTP administration inrats pretreated with SSRI. SSRI administration was performed afterthe control period and at the point where steady-state FC 5-HT levelscould be obtained (Fig. 4B). There was a small nonsignificant increasein FC 5-HT levels after SSRI administration. The mean FC 5-HT levelat 30 min after SSRI administration was 1.03 ± 0.20 pg ⁄ 10 lL, whilethe
0.46 ± 0.11 pg ⁄ 10 lL. The first blood sample was obtained duringthe control period while the second blood sample was drawn at 30 minafter SSRI pretreatment. When 5-HTP (30 mg ⁄ kg in saline solution)was given intravenously to SSRI-pretreated rats, a more rapid increasein FC 5-HT levels was observed when compared to the correspondingdata for rats without SSRI pretreatment (Fig. 2A). The third bloodsample was obtained at 30–60 min after 5-HTP administration.
Figure 4B shows individual time courses for the changes in FC
5-HT levels after 5-HTP administration in rats without SSRIpretreatment. Saline solution was administered in the same manner
Fig. 4. The effects of 5-HTP administration on individual FC 5-HT levels in
as for the SSRI-pretreated rats. When 5-HTP (30 mg ⁄ kg in saline
rats (A) with or (B) without SSRI pretreatment. Note that this experiment was
solution) was administered intravenously, we observed differences for
performed in rats that had undergone total removal of their gastrointestinal tracts
both the time course and the magnitude of the changes of the FC 5-HT
and kidneys, along with liver inactivation. (A) Time courses showing changes
levels among the five rats studied, which was similar to our findings in
for the individual FC 5-HT levels before and after 5-HTP administration in fiveSSRI pretreated rats. The arrow labelled SSRI indicates the time of pretreatment
Experiment 2. For example, rats 6 and 7 showed rapid and large
with SSRI (10 mg ⁄ kg i.v.). The arrow labelled 5-HTP indicates the time of
increases in the FC 5-HT levels after 5-HTP administration. Thus, in
5-HTP administration (30 mg ⁄ kg, i.v.). Open symbols indicate the time when
these cases blood sampling was performed at 40 min after 5-HTP
blood samples for 5-HT analysis were drawn. Note that blood samples obtained
administration. In the remaining three rats, blood sampling was not
during the control period (first blood sample) were drawn 20 min before SSRI
performed until FC 5-HT levels clearly increased. In rat 10, a blood
pretreatment. The second blood sample was obtained at 30 min after SSRIpretreatment, corresponding to 10 min before 5-HTP administration in all five
sample was obtained at 60 min after 5-HTP administration, and in rats
rats. The third blood sample was drawn at the point where we observed an
8 and 9 samples were drawn at 90 min after 5-HTP administration.
apparent increase in the FC 5-HT level after 5-HTP administration in each rat.
Figure 5A shows statistical data for changes in the FC 5-HT levels
See text for details. (B) Time courses showing changes in the individual FC
after 5-HTP administration in rats with and without SSRI pretreat-
5-HT levels before and after 5-HTP administration in the five rats that did notreceive any SSRI pretreatment (saline administration). The arrows labelled with
ment. The mean FC 5-HT level at the 5-HTP postinjection (Fig. 5A)
saline or 5-HTP indicate the times when saline or 5-HTP were administered.
represents the FC 5-HT level obtained from the last microdialysis
Open symbols indicate the times when blood samples for 5-HT analysis were
sampling point after 5-HTP administration. The mean FC 5-HT levels
obtained. The times when the first, second and third blood samples were drawn
after 5-HTP administration in rats with and without SSRI pretreatment
are the same as those described in A.
were 45.40 ± 12.75 pg ⁄ 10 lL (n ¼ 5) and 42.35 ± 12.55 pg ⁄ 10 lL(n ¼ 5), respectively. One-way anova revealed significant changes in
administration, which was in contrast to the very small change noted
the mean FC 5-HT levels after 5-HTP administration in both the rats
after the saline pretreatment (saline pretreatment, Fig. 5B). Two-way
with SSRI pretreatment (F4,2 ¼ 12.34, P < 0.01) and those without
anova revealed significant changes in the time course after 5-HTP
SSRI pretreatment (F4,2 ¼ 10.93, P < 0.01).
administration for both SSRI and saline pretreatment. As illustrated in
Figure 5B shows the statistical data for the time courses of the
Fig. 5B, the interaction effect for (SSRI + 5-HTP) administra-
changes in blood 5-HT levels after 5-HTP administration in rats with
tion · (saline + 5-HTP) administration was found to be significant
and without SSRI pretreatment. The mean blood 5-HT level before
(F1,2 ¼ 55.12, P < 0.01). Individual anova revealed a significant
SSRI pretreatment (control, Fig. 5B) was 2.02 ± 0.07 lg ⁄ mL
change in the time courses of the mean blood 5-HT level after
(n ¼ 5). There was a small decrease in the mean blood 5-HT level
(SSRI + 5-HTP) administration (F4,2 ¼ 8.34, P < 0.05) and (sal-
after SSRI pretreatment (SSRI pretreatment; Fig. 5B), whereas there
ine + 5-HTP) administration (F4,2 ¼ 69.38, P < 0.0001). With regard
was little change observed for the mean blood 5-HT level after 5-HTP
to the mean blood 5-HT level after SSRI or saline pretreatment, there
administration (5-HTP postinjection; Fig. 5B). On the other hand, the
was a significant post hoc difference noted between the control and the
mean blood 5-HT level after 5-HTP administration in rats without
SSRI ⁄ saline pretreatments (P < 0.05) only in the SSRI-pretreated
SSRI pretreatment (Fig. 5B) showed a marked increase after 5-HTP
rats. As for the mean blood 5-HT level after 5-HTP administration,
ª The Authors (2008). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing Ltd
European Journal of Neuroscience, 27, 2466–2472
that all neurotransmitters are retained in the brain. However, recentin vitro studies (Brust et al., 2000; Wakayama et al., 2002) haverevealed the presence of 5-HT transporter mRNA in vascularendothelial cells. This indicates that the BBB may act as an effluxtransport system for 5-HT. Based on this new data, we conducted thepresent in vivo functional study. As a result, we found that indeed5-HT can cross from the brain into the circulating blood via the BBB.
The second reason that is cited as to why 5-HT cannot cross the
BBB is related to the 5-HT content that is found within the organs. Ithas been reported that > 90% of the total 5-HT content in the wholebody is distributed within the gastrointestinal tract (West, 1958;Gaginella, 1995), and that only a very small percentage of the total5-HT content is found within the brain. Therefore, it has been believedthat the augmented ECF 5-HT in the brain does not contribute to anysignificant changes in the 5-HT levels within the circulating blood. Therefore, we administered 5-HTP intravenously in rats that hadundergone the abdominal operation in an attempt to elevate brain5-HT alone. As a result, we found that whole-blood 5-HT levelssignificantly increased whenever brain ECF 5-HT levels were elevatedby the 5-HTP administration in the rats undergoing the abdominaloperation. Therefore, it is reasonable to hypothesize that theaugmented brain ECF 5-HT does contribute to a significant changein 5-HT levels within the circulating blood. In other words, augmentedbrain ECF 5-HT can translocate from the brain into the blood via theBBB.
This hypothesis may be further supported by the data concerning
regional differences of the 5-HTP decarboxylase, which is the enzymeresponsible for metabolizing 5-HTP to 5-HT. A study by West (1958)demonstrated that there was high 5-HTP decarboxylase activity in thekidneys (188 lg ⁄ tissue) and the liver (125 lg ⁄ tissue), with only avery low activity noted in the gastrointestinal tract (1–2 lg ⁄ tissue). The brain exhibited moderate activity for 5-HTP decarboxylase(32 lg ⁄ tissue). In addition, 5-HT that is produced by the gastrointes-
Fig. 5. The effects of 5-HTP administration on (A) FC 5-HT levels and
tinal tract is thought to be metabolized by monoamine oxidase in the
(B) blood 5-HT levels in rats with SSRI or saline pretreatment. All rats
liver, as it circulates through the portal vein (Gillis, 1985). Therefore,
underwent a surgical procedure to totally remove their gastrointestinal tractsand kidneys, along with liver inactivation. Arrows labelled SSRI or saline
it is unlikely that the gastrointestinal tract would make any significant
indicate the times of the SSRI (10 mg ⁄ kg i.v) or saline pretreatments. Arrows
contribution towards increasing blood 5-HT when 5-HTP is admin-
labelled 5-HTP indicate the times of the 5-HTP (30 mg ⁄ kg i.v) administrations.
s, Data obtained in rats with SSRI pretreatment (n ¼ 5); d, data points from
With regard to intrarenal formation of 5-HT by renal decarboxylase,
rats without SSRI pretreatment (saline administration, n ¼ 5). Data areexpressed as the mean ± SE. #P < 0.05, ##P < 0.01, ####P < 0.0001 vs.
Stier & Itskovitz, 1985) demonstrated that administration of 5-HTP
control. **P < 0.01, SSRI + 5-HTP vs. Saline + 5-HTP.
resulted in an increase in urinary 5-HT without a concomitant increasein plasma 5-HT in the rat. Based on this result, it is reasonable to
there was a significant post hoc difference observed between the
speculate that intrarenal formation of 5-HT may not contribute to an
control and the 5-HTP postinjection (P < 0.0001) in the rats that were
increase in 5-HT in whole blood after 5-HTP administration.
Based on data by West (1958), the whole organs that exhibit 5-HTP
There was also a significant post hoc difference noted in the blood
decarboxylase activity include the kidneys, the liver, the gastrointes-
5-HT level for the 5-HTP postinjection between the SSRI- and saline-
tinal tract, the brain and the skin. After 5-HTP administration in rats
that underwent resection of their gastrointestinal tracts and kidneysalong with liver inactivation, the skin in addition to the brain were thetwo organs found to be capable of augmenting the whole-blood 5-HT
levels. However, skin has been demonstrated to show the lowest
The present study revealed that whole-blood 5-HT levels exhibited
amount of enzyme activity (1 lg ⁄ tissue). Thus, it is less likely that
significant augmentation when brain 5-HT levels were elevated after
5-HT in the skin contributes to any significant augmentation of 5-HT
5-HTP administration in rats that had undergone total removal of their
in the whole blood after 5-HTP administration, even though we can
gastrointestinal tracts and kidneys, along with liver inactivation.
not completely rule out this possibility.
This result implies that 5-HT may cross the brain into the circulating
The other new finding of the present study is that SSRI pretreatment
blood via the BBB. However, there are two major reasons that may be
in rats lacking functional kidneys, gastrointestinal tract and liver
cited as evidence as to why 5-HT cannot possibly cross from the brain
abolished elevation of whole-blood 5-HT levels that was induced by
into the circulating blood via the BBB.
5-HTP administration, even though the brain 5-HT levels remained
First, the BBB is formed by tight junctions of the brain capillary
increased. These results suggest that the 5-HT transporters that are
endothelial cells. The role of these junctions is to prevent neurotrans-
located on the brain endothelial cells play the inevitable role
mitters including 5-HT from crossing the junction and thus ensuring
associated with crossing from the brain via the BBB into the
ª The Authors (2008). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 27, 2466–2472
circulating blood. As discussed earlier, the in vitro study by Brust
Brust, P., Friedrich, A., Krizbai, I.A., Bergmann, R., Roux, F., Ganapathy, V. &
et al. (2000) revealed the presence of a 5-HT transporter mRNA in the
Johannsen, B. (2000) Functional expression of the serotonin transporter inimmortalized rat brain microvessel endothelial cells. J. Neurochem., 74,
brain endothelium, indicating that cerebral endothelial cells are able to
actively participate in the removal of the released 5-HT within the
Fuxe, K. & Agnati, L.F. (1991) Two principal modes of electrochemical
brain. This possibility was functionally proven in the present in vivo
communication in the brain: volume versus wiring transmission. In Fuxe, K.
physiological study that used the SSRI. Thus, we hypothesized that
& Agnati, L.F. (eds), Volume Transmission in the Brain: Novel Mechanisms
the 5-HT transporters located on the brain endothelial cells may act as
for Neural Transmission. Raven Press, New York, pp. 1–9.
Fuxe, K., Dahlstro¨m, A., Ho¨istad, M., Marcellino, D., Jansson, A., Rivera, A.,
the efflux transport system for the 5-HT that crosses from the brain
Diaz-Cabiale, Z., Jacobsen, K., Tinner-Staines, B., Hagman, B., Leo, G.,
Staines, W., Guidolin, D., Kehr, J., Genedani, S., Belluardo, N. & Agnati,
What physiological role does the 5-HT transporter located on the
L.F. (2007) From the Golgi–Cajal mapping to the transmitter-based
brain endothelium actually play? It has been established that the 5-HT
characterization of the neuronal networks leading to two modes of braincommunication: Wiring and volume transmission. Brain Res. Rev., 55, 17–
transporter is present not only in the synaptic terminals of 5-HT
neurons but also in the brain endothelial cells (Brust et al., 2000;
Gaginella, T.S. (1995) Serotonin in the intestinal tract: a synopsis. In Gaginella,
Wakayama et al., 2002). Therefore, we can hypothesize that both 5-HT
T.S. & Galligan, J.J. (eds), Serotonin and Gastrointestinal Function. CRC
transporters play a significant role in 5-HT homeostasis within the
brain. There is no doubt that 5-HT transporters located on the terminals
Gartside, S.E., Cowen, P.J. & Sharp, T. (1992) Effect of 5-hydroxy-L-
tryptophan on the release of 5-HT in rat hypothalamus in vivo as measured
of 5-HT neurons play an inevitable role in 5-HT homeostasis within the
by microdialysis. Neuropharmacology, 31, 9–14.
synaptic cleft. On the other hand, it has been reported that 5-HT’s role
Gillis, C.N. (1985) Peripheral metabolism of serotonin. In Vanhoutte, P.M.
is more in line with volume transmission than classical neurotrans-
(ed.), Serotonin and the Cardiovascular System. Raven Press, New York, pp.
mission (Fuxe & Agnati, 1991; Hornung, 2003; Fuxe et al., 2007).
Hornung, J.P. (2003) The human raphe nuclei and the serotonergic system.
5-HT released from the axonal terminals and varicosities diffuses over
a long distance to act on other neurons. In this particular case, the
Jacobs, B.L. & Azmitia, E.C. (1992) Structure and function of the brain
released 5-HT could be detected as a change in ECF 5-HT in the brain.
serotonin system. Physiol. Rev., 72, 165–229.
Augmented ECF 5-HT within the brain may be drained away via the
Kremer, H.P., Goekoop, J.G. & Van Kempen, G.M. (1990) Clinical use of the
endothelial 5-HT transporter that is located in the nearby endothelial
determination of serotonin in whole blood. J. Clin. Psychopharmacol., 10,83–87.
cells, thereby maintaining 5-HT homeostasis in the brain. In other
Lo¨scher, W., Pagliusi, S.R. & Mu¨ller, F. (1984) L-5-hydroxytryptophan.
words, the physiological role of the BBB is not only to act as a barrier
Correlation between anticonvulsant effect and increases in levels of
but also to play a role as a regulatory interface for brain ECF 5-HT.
5-hydroxyindoles in plasma and brain. Neuropharmacology, 23, 1041–
In summary, increased brain ECF 5-HT is removed from the brain
Okada, F., Saito, Y., Fujieda, T. & Yamashita, I. (1972) Monoamine changes
into the circulating blood via the 5-HT transporter system located on
in the brain of rats injected with L-5-hydroxytryptophan. Nature, 238, 355–
Ortiz, J., Artigas, F. & Gelpı´, E. (1988) Serotonergic status in human blood.
Paxinos, G. & Watson, C. (1986) The Rat Brain in Stereotaxic Coordinates,
5-HT, 5-hydroxytryptamine, serotonin; 5-HTP, 5-hydroxytryptophan; BBB,
Penalva, R.G., Lancel, M., Flachskamm, C., Reul, J.M., Holsboer, F. &
blood–brain barrier; ECF, extracellular fluid; FC, frontal cortex; HPLC, high-
Linthorst, A.C. (2003) Effect of sleep and sleep deprivation on serotonergic
performance liquid chromatography; SSRI, selective serotonin reuptake
neurotransmission in the hippocampus: a combined in vivo microdialy-
sis ⁄ EEG study in rats. Eur. J. Neurosci., 17, 1896–1906.
Stier, C.T. Jr & Itskovitz, H.D. (1985) Formation of serotonin by rat kidneys
in vivo. Proc. Soc. Exp. Biol. Med., 180, 550–557.
Wakayama, K., Ohtsuki, S., Takanaga, H., Hosoya, K. & Terasaki, T. (2002)
Localization of norepinephrine and serotonin transporter in mouse brain
Artigas, F., Sarrias, M.J., Martı´nez, E. & Gelpı´, E. (1985) Serotonin in body
capillary endothelial cells. Neurosci. Res., 44, 173–180.
fluids: characterization of human plasmatic and cerebrospinal fluid pools by
West, G.B. (1958) Studies on 5-hydroxytryptamine and 5-hydroxytryptophan.
means of a new HPLC method. Life Sci., 37, 441–447.
J. Pharm. Pharmacol., 10 (Suppl.), 92–97.
ª The Authors (2008). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing Ltd
European Journal of Neuroscience, 27, 2466–2472
Module de Base 4 Pharmacologie générale, bases de thérapeutique générale. Devenir des médicaments dans l’organisme(Pharmacocinétique). ELIMINATION Objectifs Interpréter correctement les informations pharmacocinétiques suivantes, en relation avec les schémas posologiques et les précautions d'utilisation . Fraction bio-disponible F d'une dose orale D pharmacothér
Deutsche Gesellschaft für Gynäkologie und Geburtshilfe e.V. Leitlinien, Empfehlungen, Stellungnahmen 3.4.5. Schwangerenbetreuung und Geburtsleitung bei Zustand nach Kaiserschnitt Deutsche Gesellschaft für Gynäkologie und Geburtshilfe (DGGG), Board für Pränatal- und Geburtsmedizin, Arbeitsgemeinschaft Materno-fetale Medizin, Deutsche Gesellschaft für Perinatale Medizin (DGPM)
 served as the control animals. Following the injection of 5-HTP or
saline solution, samples were obtained from the FC every 10 min for
Experiment 1: effect of 5-HTP administration on the frontal
cortex (FC) and blood 5-HT levels in intact rats
Blood sampling was performed three times in the six rats given
5-HTP. The first blood sample was taken prior to the injection of
The first experiment was performed in intact rats that did not undergo
5-HTP (preinjection sample). The second blood sample was obtained
the abdominal operation. Figure 1A shows the statistical data for the
when the brain 5-HT level had increased to more than two times
time course of the changes in FC 5-HT levels before and after 5-HTP
higher than the preinjection level, which occurred at 50–70 min after
administration. The mean preinjection 5-HT level in FC was
5-HTP administration. The third blood sample was taken when the
0.39 ± 0.04 pg ⁄ 10 lL (n ¼ 5). When 5-HTP was administered
brain 5-HT level further increased, which occurred at 10–30 min after
intravenously, there was a gradual increase in FC 5-HT levels.
served as the control animals. Following the injection of 5-HTP or
saline solution, samples were obtained from the FC every 10 min for
Experiment 1: effect of 5-HTP administration on the frontal
cortex (FC) and blood 5-HT levels in intact rats
Blood sampling was performed three times in the six rats given
5-HTP. The first blood sample was taken prior to the injection of
The first experiment was performed in intact rats that did not undergo
5-HTP (preinjection sample). The second blood sample was obtained
the abdominal operation. Figure 1A shows the statistical data for the
when the brain 5-HT level had increased to more than two times
time course of the changes in FC 5-HT levels before and after 5-HTP
higher than the preinjection level, which occurred at 50–70 min after
administration. The mean preinjection 5-HT level in FC was
5-HTP administration. The third blood sample was taken when the
0.39 ± 0.04 pg ⁄ 10 lL (n ¼ 5). When 5-HTP was administered
brain 5-HT level further increased, which occurred at 10–30 min after
intravenously, there was a gradual increase in FC 5-HT levels.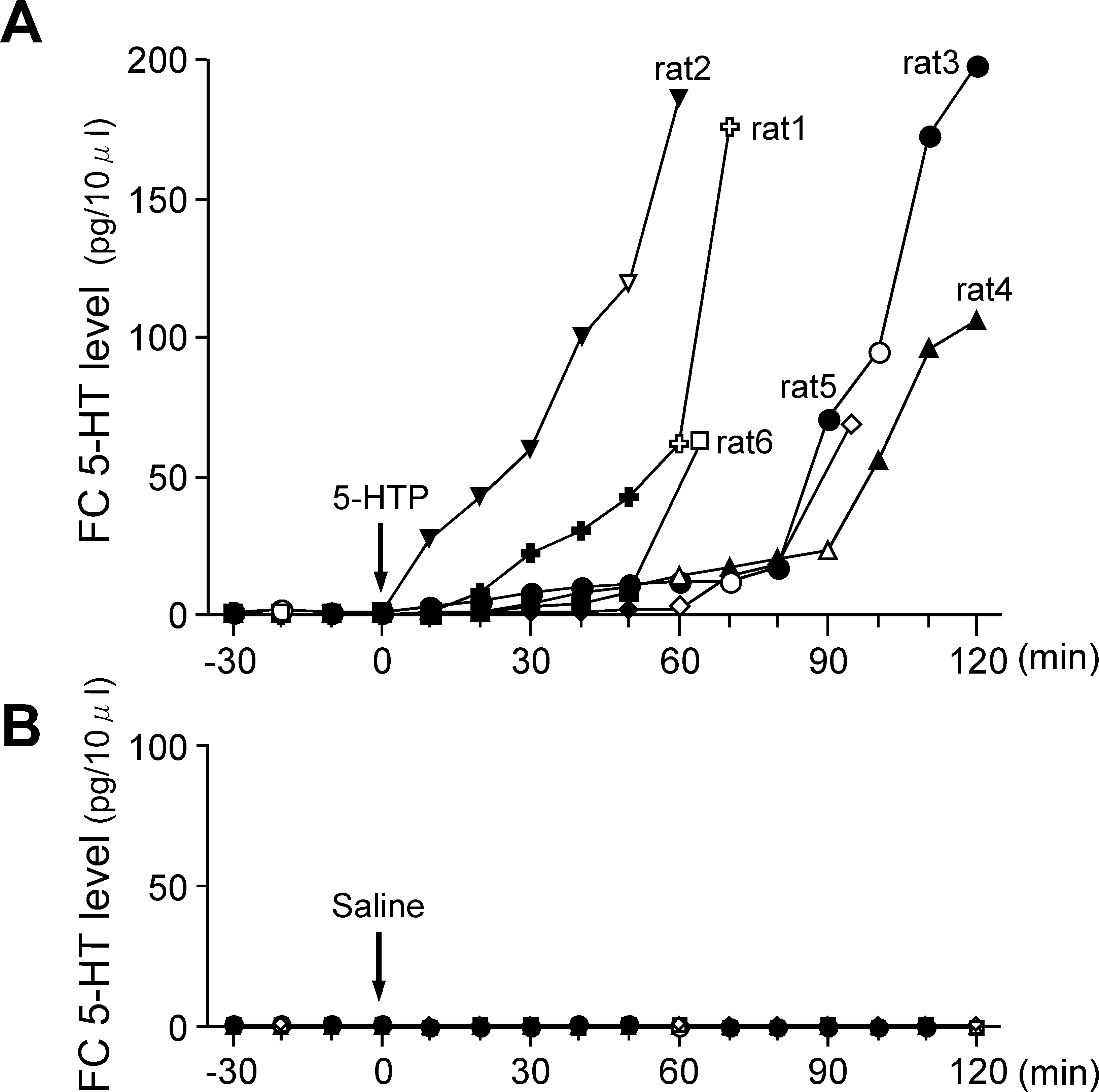
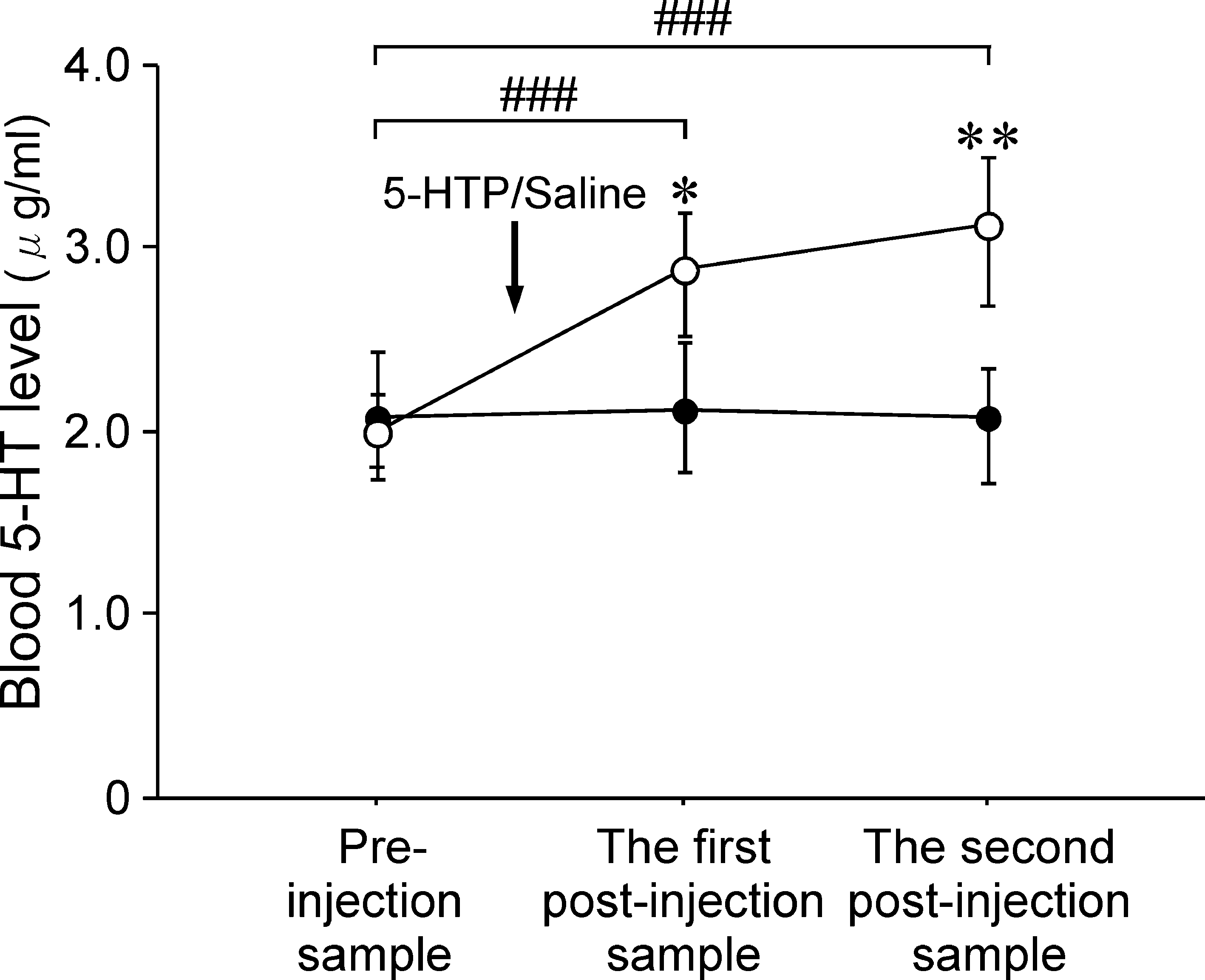 Figure 1B shows the time course for the changes in blood 5-HT
at 60 min after 5-HTP administration. On the other hand, rat 4
levels. The mean blood 5-HT level before 5-HTP administration was
expressed a slow and small increase in the FC 5-HT level after 5-HTP
1.95 ± 0.10 lg ⁄ mL. A one-way anova revealed significant changes
administration, with a FC 5-HT level of 14.1 pg ⁄ 10 lL at 60 min and
in the time course for the mean blood 5-HT level after 5-HTP
a peak FC 5-HT level of 105.8 pg ⁄ 10 lL at 120 min after 5-HTP
administration (F4,3 ¼ 111.5, P < 0.0001). There was a significant
administration. As the time courses for the changes in FC 5-HT levels
post hoc difference between before and 60 min (P < 0.0001),
differed among the six rats after 5-HTP administration, we obtained
120 min (P < 0.001) and 180 min (P < 0.001) after 5-HTP admin-
the first postinjection blood sample at a time when the FC 5-HT level
istration. The maximum blood 5-HT level observed at 60 min after the
was more than two times higher than the preinjection level. Thus, the
5-HTP injection was 4.40 ± 0.14 lg ⁄ mL, which was approximately
first postinjection blood samples (open symbol for each response
two times higher than the preinjection blood 5-HT level. The blood
curve) were drawn between 50 and 70 min after 5-HTP administration
5-HT levels continued to remain high until 180 min after 5-HTP
in this experiment. The second postinjection samples were taken when
administration, although FC 5-HT levels showed an apparent decrease
the FC 5-HT level further increased, which occurred at 10–30 min
after the first postinjection blood samples were drawn.
Figure 1B shows the time course for the changes in blood 5-HT
at 60 min after 5-HTP administration. On the other hand, rat 4
levels. The mean blood 5-HT level before 5-HTP administration was
expressed a slow and small increase in the FC 5-HT level after 5-HTP
1.95 ± 0.10 lg ⁄ mL. A one-way anova revealed significant changes
administration, with a FC 5-HT level of 14.1 pg ⁄ 10 lL at 60 min and
in the time course for the mean blood 5-HT level after 5-HTP
a peak FC 5-HT level of 105.8 pg ⁄ 10 lL at 120 min after 5-HTP
administration (F4,3 ¼ 111.5, P < 0.0001). There was a significant
administration. As the time courses for the changes in FC 5-HT levels
post hoc difference between before and 60 min (P < 0.0001),
differed among the six rats after 5-HTP administration, we obtained
120 min (P < 0.001) and 180 min (P < 0.001) after 5-HTP admin-
the first postinjection blood sample at a time when the FC 5-HT level
istration. The maximum blood 5-HT level observed at 60 min after the
was more than two times higher than the preinjection level. Thus, the
5-HTP injection was 4.40 ± 0.14 lg ⁄ mL, which was approximately
first postinjection blood samples (open symbol for each response
two times higher than the preinjection blood 5-HT level. The blood
curve) were drawn between 50 and 70 min after 5-HTP administration
5-HT levels continued to remain high until 180 min after 5-HTP
in this experiment. The second postinjection samples were taken when
administration, although FC 5-HT levels showed an apparent decrease
the FC 5-HT level further increased, which occurred at 10–30 min
after the first postinjection blood samples were drawn.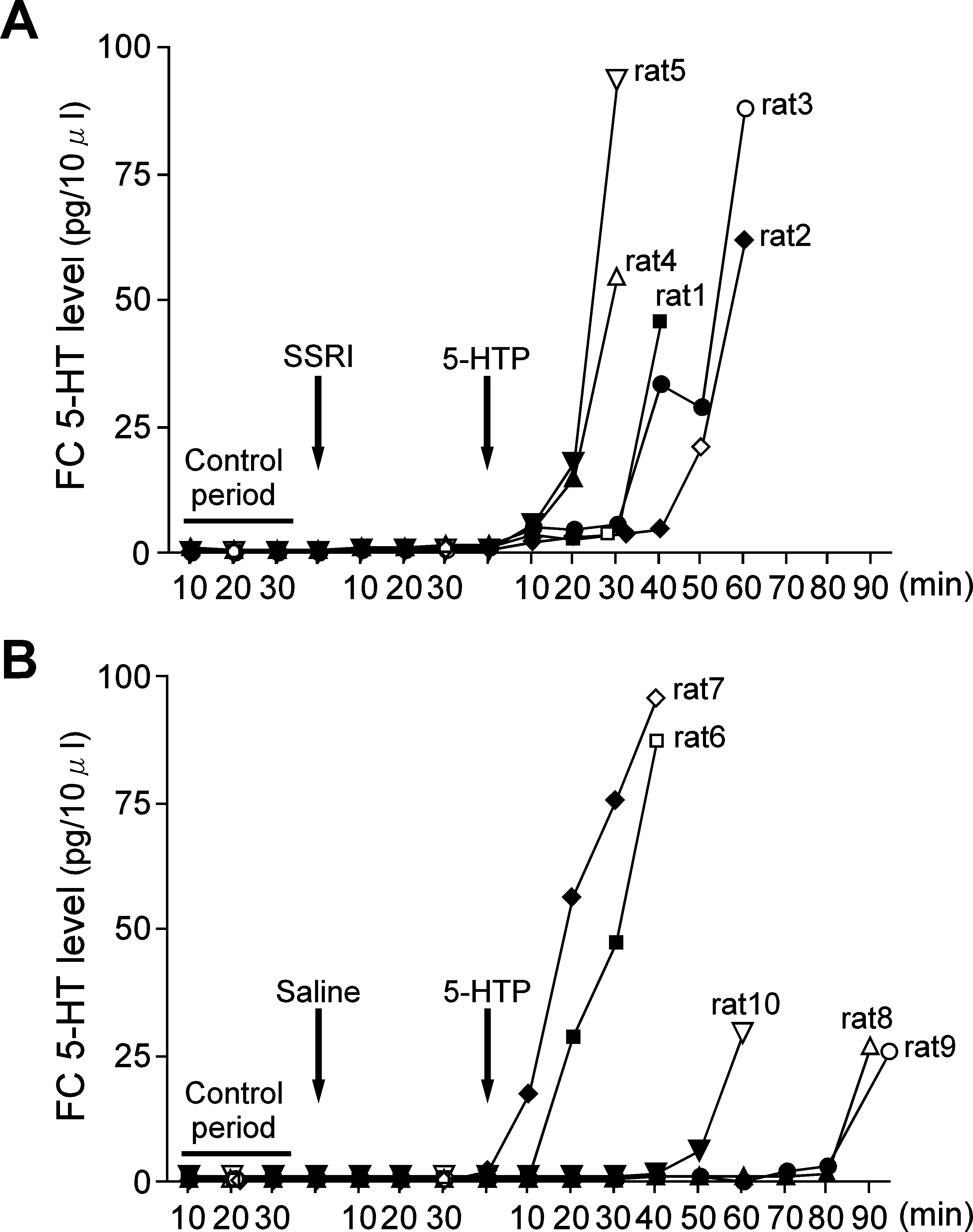 significant post hoc difference observed between the preinjectionsample and the first (P < 0.001) and second (P < 0.001) postinjectionsamples. There was also a significant post hoc difference between5-HTP administration and saline administration for both the first(P < 0.05) and second (P < 0.01) postinjection samples.
significant post hoc difference observed between the preinjectionsample and the first (P < 0.001) and second (P < 0.001) postinjectionsamples. There was also a significant post hoc difference between5-HTP administration and saline administration for both the first(P < 0.05) and second (P < 0.01) postinjection samples.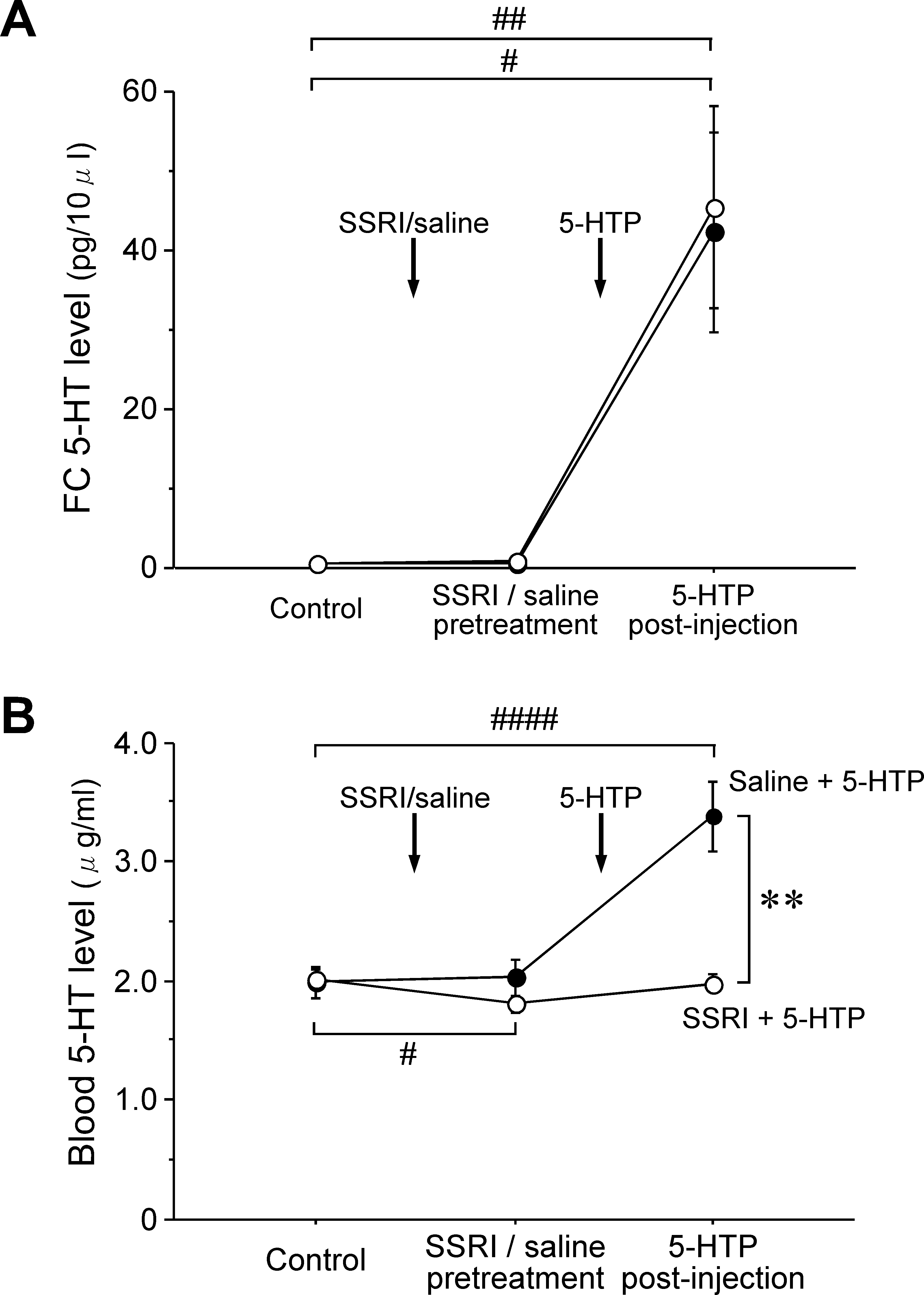 that all neurotransmitters are retained in the brain. However, recentin vitro studies (Brust et al., 2000; Wakayama et al., 2002) haverevealed the presence of 5-HT transporter mRNA in vascularendothelial cells. This indicates that the BBB may act as an effluxtransport system for 5-HT. Based on this new data, we conducted thepresent in vivo functional study. As a result, we found that indeed5-HT can cross from the brain into the circulating blood via the BBB.
that all neurotransmitters are retained in the brain. However, recentin vitro studies (Brust et al., 2000; Wakayama et al., 2002) haverevealed the presence of 5-HT transporter mRNA in vascularendothelial cells. This indicates that the BBB may act as an effluxtransport system for 5-HT. Based on this new data, we conducted thepresent in vivo functional study. As a result, we found that indeed5-HT can cross from the brain into the circulating blood via the BBB.