Tadalafil appartiene alla classe degli inibitori selettivi della fosfodiesterasi di tipo 5, con un profilo farmacocinetico caratterizzato da un’emivita terminale di circa diciotto ore. Dopo somministrazione orale viene assorbito rapidamente e raggiunge concentrazioni plasmatiche massime in due ore. La biotrasformazione avviene principalmente tramite CYP3A4 con formazione di metaboliti inattivi, escreti in prevalenza con le feci. L’elevato legame alle proteine plasmatiche (>90%) assicura una distribuzione stabile. Nei confronti delle altre molecole della stessa classe, cialis compresse italia è noto per la durata prolungata dell’attività farmacologica.
An extremely low frequency magnetic field attenuates insulin secretion from the insulinoma cell line, rin-m
An Extremely Low Frequency Magnetic Field
Tomonori Sakurai,1 Akira Satake,1 Shoichiro Sumi,1 Kazutomo Inoue,1 and Junji Miyakoshi2*
1Department of Organ Reconstruction, Institute for Frontier Medical Sciences,
2Department of Radiological Technology, School of Health Sciences,
Faculty of Medicine, Hirosaki University, Hirosaki, Japan
In this study, we investigated the effects of exposure to an extremely low frequency magnetic field(ELFMF) on hormone secretion from an islet derived insulinoma cell line, RIN-m. We stimulatedRIN-m cells to secrete insulin under exposure to an ELFMF, using our established system for theexposure of cultured cells to an ELFMF at 5 mT and 60 Hz, or under sham exposure conditions for 1 hand observed the effects. In the presence of a depolarizing concentration of potassium (45 mM KCl),exposure to ELFMF significantly attenuated insulin release from RIN-m cells, compared to shamexposed cells. Treatment with nifedipine reduced the difference in insulin secretion betweencells exposed to an ELFMF and sham exposed cells. The expression of mRNA encoding synaptosomalassociated protein of 25 kDa (SNAP-25) and synaptotagmin 1, which play a role in exocytosis inhormone secretion and influx of calcium ions, decreased with exposure to an ELFMF in the presenceof 45 mM KCl. These results suggest that exposure to ELFMF attenuates insulin secretion fromRIN-m cells by affecting calcium influx through calcium channels. Bioelectromagnetics 25:160–166,2004.
Key words: 5 mT and 60 Hz; insulinoma cell line; KCl stimulation; calcium channels;
synaptosomal associated protein; synaptotagmin 1
ion efflux, and insulin secretion during glucose stimu-lation were reduced when isolated rabbit islets were
The possible health effects of exposure to ex-
exposed to low frequency pulsed magnetic fields.
tremely low frequency magnetic fields (ELFMFs) have
Hayek et al. [1984] reported that exposure to low
become a considerable public concern. Several epide-
intensity homogeneous magnetic fields inhibited insu-
miological studies have shown an association between
lin release from isolated newborn rat islets stimulated
exposure to ELFMF and elevated risk in children and
by high glucose concentration (16.7 mM) and amino-
occupationally exposed adults [Savitz and Loomis,
phylline (10 mM). Recently, Laitl-Kobierska et al.
1995]. Whether exposure to magnetic fields causes
[2002] reported that long term exposure of rats to
significant cellular stress remains a contentious issue in
ELFMF led to increased synthesis and secretion of
Pancreatic islets play a fundamental role in re-
gulating the blood glucose levels of the body through
Grant sponsor: Research for the Future Program, Japan Society for
the secretion of hormones such as insulin, glucagon,
the Promotion of Science; Grant sponsor: Ministry of Education
somatostatin, and pancreatic polypeptide. The release
(for Scientific Research S); Grant number: 13854020.
of insufficient amounts of these hormones is the basis of
*Correspondence to: Prof. Junji Miyakoshi, Department of
various forms of diabetes. Therefore, it is important to
Radiological Technology, School of Health Sciences, Faculty of
assess the effects of exposure to ELFMF on pancreatic
Medicine, Hirosaki University, 66-1 Hon-cho, Hirosaki, 036-8564,
Japan. E-mail: [email protected]
Studies evaluating the influence of exposure to
ELFMF on pancreatic islet function are scarce, and it is
Received for review 2 May 2003; Final revision received 4 August2003
often difficult to compare the existing studies because ofthe different research methods used. Previously, Joelly
et al. [1983] reported that calcium ion content, calcium
Published online in Wiley InterScience (www.interscience.wiley.com).
insulin. Consequently, an association between mag-
netic field exposure and pancreatic islet function has not
was removed, and the cells were washed twice with 2-
[4-(2-hydroxyethyl)-1-piperazinyl]ethanesulfonic acid
One important approach to overcome the pro-
(HEPES)-buffered Krebs–Ringer solution (119 mM
blems of using islets is the use of insulinoma cell lines.
NaCl, 4.74 mM KCl, 2.54 mM CaCl2, 1.19 mM
RIN-m cells are derived from X-ray radiation induced
rat insulinoma [Chick et al., 1977; Gazdar et al., 1980]
10 mM HEPES at pH 7.4) containing 0.2% bovine
and have been used to investigate the mechanism of
serum albumin (BSA), and 3.3 mM D-glucose. The cells
insulin secretion [Yada et al., 1989].
were then preincubated in the same buffer at 37 8C
In this study, we have investigated the effects of
for 30 min. The buffer was then removed, and 0.8 ml
exposure to ELFMF on insulin release by the insuli-
of basal incubation buffer (HEPES-buffered Krebs–
noma cell line, RIN-m, using our previously manufac-
Ringer solution containing 0.2% BSA and 3.3 mM
tured equipment to expose cultured cells to an ELFMF
D-glucose) was added and the cells were incubated at
37 8C for 1 h. After the basal incubation period, wellswere divided into two groups; one is stimulated underexposure to an ELFMF (Fig. 1, Group A), and the other
is stimulated under sham exposure condition (Fig. 1,
The stimulation was performed as follows: the
ELFMF exposure, a sinusoidal magnetic field at a
cells were incubated at 37 8C for 1 h in 0.8 ml of
frequency of 60 Hz, 5 mT, was performed using a pre-
stimulation buffer, which consisted of HEPES buffered
viously described magnetic field exposure apparatus
Krebs–Ringer solution containing 0.2% BSA, D-glucose,
[Miyakoshi et al., 1996; Ding et al., 2000]. The distri-
and various reagents, as indicated in the figure legends.
bution of the magnetic density was measured using a
At the end of the basal incubation period and stimu-
Gauss meter (Model 3251, Yokogawa Electrical Co.,
lation period, an aliquot of the incubation buffer in each
Ltd., Tokyo). Briefly, the ELFMF exposure system
well was collected and stored at À20 8C until the insulin
consists of a magnetic field generator that uses
measurement was performed. The insulin concentration
Helmholtz coils built into a CO2 incubator, a transfor-
of the samples was measured by enzyme linked im-
mer and a thermocontroller. The direction of the field is
munosorbent assay (ELISA) using a rat insulin ELISA
vertical. The atmosphere in the incubator is maintained
kit (Sibayagi Co., Gunma, Japan). Insulin secretion
with humidified 95% air plus 5% CO2. The temperature
from RIN-m cells stimulated various regents were
in the exposure space, which is monitored by thermo-
analyzed using the ratio of insulin secretion during the
couple sensor probes, is maintained at 37 Æ 0.2 8C. The
stimulation period/insulin secretion during basal incu-
measured 60 Hz ELFMF exposure during the sham
exposure was <0.5 mT. Static magnetic fields other thangeomagnetism were undetectable (<0.1 mT).
After insulin secretion tests were performed using
6-well culture plates, the cells were scraped and the
RIN-m cells (obtained from the Dainippon Phar-
total RNA was prepared using an ISOGEN isolation kit
maceutical Co., Osaka, Japan) were cultured in RPMI-
(Nippon Gene Co., Toyama, Japan). Briefly, the cells
1640 medium supplemented with 10% fetal bovine
were homogenized in 1 ml of ISOGEN reagent, and
serum at 37 8C in humidified 95% air plus 5% CO2. For
0.1 ml of chloroform was added to this mixture. The
each experiment, a new vial of frozen cells was thawed,
mixture was centrifuged at 15,000g for 15 min at 4 8C,
seeded at a density of 1 Â 105 cells/cm2 on 12- or 6-well
and the resulting aqueous phase was transferred to
cell culture plates and cultured. Cells were used from
0.5 ml of isopropyl alcohol. The resulting precipitate
was collected by centrifugation at 15,000g for 10 min at4 8C, and the RNA pellet obtained was washed with
75% ethanol, and then dissolved in diethyl pyrocarbo-
Cells were plated on 12-well culture plates at
nate-treated water. The amount of RNA was measured
a density of 3.5 Â 105 cells/well. Insulin secretion tests
using an ultraviolet-visible spectrometer at 260 nm, and
were performed 4 days after plating, when the cells were
the purity of the RNA obtained was determined from the
80–90% confluent. The culture medium was changed
absorbance ratio at 260/280 nm. The ratio at >1.8 was
to fresh medium 16 h before the insulin secretion tests,
used following reverse transcriptase-polymerase chain
and the tests were performed as shown in Figure 1.
Fig. 1. The outline of insulin secretion tests. Preincubation and basal incubation were performedwithoutexposure to anextremelylow frequencymagnetic field (ELFMF).After wellswere dividedintotwo groups, one group is stimulated under exposure to an ELFMF (Group A), and the other group isstimulatedunder sham exposure (Group B). At the end ofthe basalincubation period and stimulationperiod, an aliquot of the incubation buffer in each well was collected, and the insulin concentration ofthe samples was measured by enzyme linked immunosorbent assay (ELISA).
in a thermal cycler at 95 8C for 45 s, at 59 8C for 45 s,
cDNA synthesis was performed using a (dT)
and at 72 8C for 90 s. b-Actin expression was used to
oligo primer and ThermoScript RT (Invitrogen, Carls-
normalize the input template cDNA in a semi-quanti-
bad, CA), according to the manufacturer’s instructions.
tative PCR reaction. Serial half dilutions of cDNA were
PCR was performed using Taq polymerase (TaKaRa
amplified to ensure analysis of products in the linear
Co, Shiga, Japan). PCR mixtures contained cDNA, 1Â
PCR buffer, 2.5 mM MgCl2, 400 mM dNTP, 2.5 U/50 ml
of Taq polymerase, and 1 mM sense and antisenseprimers. The primers were as follows, insulin 1: forward
50-ATGGCCCTGTGGATGCGCTT-30, reverse 50-TA-
agarose gel with 0.1 mg/ml ethidium bromide, and the
GTTGCAGTAGTTCTCCAGCT-30, insulin 2: forward
quantification of the PCR products was performed by
densitometry of the band intensity using a Kodak Digi-
tal Science IS 440 CF System and 1D Image Analysis
associated protein of 25 kDa (SNAP-25): forward 50-
Software ver. 3.5 (Eastman Kodak Co., Rochester, NY).
GGTTCCTTAACTAAGCACCACTGACTT-30, reverse50-TTTCCCGGGCATCGTTTGTTACC-30,
tagmin 1: forward 50-ATGGCTGTGTATGACTTTGA-
Statistical comparisons were performed by analy-
sis of variance and, when appropriate, using the Stu-
GTCGTT-30, b-actin: forward 50-ATGGTGGGTATG-
dent’s t-test. Experimental results were presented as the
mean Æ SE, and studies were repeated at least three
CTCTCAGTC-30. The reaction mixture was incubated
Insulin Secretion From RIN-m CellsUnder Sham Conditions
Under sham conditions, incubation with stimula-
tion buffer containing a high concentration of potas-sium (45 mM KCl) caused insulin release to increaseapproximately 14 fold, compared to the basal incuba-tion conditions. In contrast, both a normal (3.3 mM)and a high (16.7 mM) concentration of D-glucose didnot increase insulin release, compared to the basal in-cubation conditions. Phorbol-12-myristate-13-acetate(PMA) also increased insulin release from RIN-m cellsby approximately 15 fold (Table 1). These results arein agreement with previous reports describing RIN-mcells and their subcloned cell line, RINm5F [Praz et al.,1983; Bhathena et al., 1984; Yada et al., 1989].
Insulin secretion from RIN-m cells in the stimula-
tion buffer containing a normal concentration of D-glucose did not increase under exposure to an ELFMF,
Fig. 2. Relative insulin release from RIN-m cells under exposure
compared to the sham exposed cells. On the other hand,
to an ELFMF or under sham conditions, measured after1h incuba-tion with 3.3 mM
in the presence of 45 mM KCl, exposure to ELFMF
D-glucose, or 3.3 mM D-glucose plus 45 mM KCl,
or 3.3 mM D-glucose plus 1 mM phorbol-12-myristate-13-acetate
significantly attenuated insulin release from RIN-m
(PMA). Data represent the mean Æ SE (3.3 mM D-glucose, n ¼ 3;
cells by approximately 30%, compared to sham exposed
45 mM KCl and PMA, n ¼ 5). **P < .01.
cells. When PMA was included in the stimulation buf-fer, insulin release was slightly, but not significantly,enhanced by exposure to ELFMF (Fig. 2). These resultssuggest that exposure to an ELFMF in the presence of
concentrations of the calcium channel blocker, nifedi-
chemical reagents affects insulin secretion, but that
pine. Under exposure to an ELFMF and under sham
exposure to ELFMF alone is insufficient to stimulate
conditions, insulin release decreased in a dose-depen-
insulin secretion. These findings are consistent with our
dent manner. In the presence of low concentrations of
previous reports showing that exposure to an ELFMF
nifedipine (5, 50, or 500 nM), insulin secretion was
in combination with chemical reagents enhanced the
significantly attenuated under exposure to an ELFMF,
effects of exposure to ELFMF on cells [Miyakoshi et al.,
compared to sham exposed cells. In contrast, there was
no difference in insulin release between cells exposed toan ELFMF and those under sham exposure at high
Effects of Nifedipine Treatment on Insulin
concentrations of nifedipine (5 or 50 mM). The amount
of insulin secretion in the presence of 5 nM nifedipine
Insulin release from RIN-m cells was assessed in
under sham conditions was approximately equal to that
the presence of 45 mM KCl after treatment with various
in the absence of nifedipine under exposure to ELFMF(Fig. 3). These results suggest that ELFMF attenuatedinsulin secretion from RIN-m cells is related to an effect
TABLE 1. Insulin Secretion From RIN-m Cells in Response to
The effects of an ELFMF on mRNA expression in
RIN-m cells were investigated using semi-quantitative
RT-PCR. The fragments amplified for insulin 1, insulin
2, SNAP-25, synaptotagmin 1, and b-actin had the ex-
pected sizes (331, 333, 493, 456, and 500 base pairs
Fig. 3. Nifedipine inhibition of insulin release from RIN-m cells sti-mulated by 3.3 mM D-glucose and 45 mMKClunderexposure to anELFMF or under sham conditions.To study the effect of nifedipine,cells were incubated with the indicated concentrations of nifedi-pine for 30 min, and then incubated for 1 h with stimulation buffercontaining 3.3 mM D-glucose and 45 mM KCl. Each point repre-
Fig. 4. A: Reverse transcription-polymerase chain reaction (RT-
sents the mean Æ SE (50 mM nifedipine, n ¼ 3; other concentra-
PCR) of insulin 1, insulin 2, synaptosomal associated protein of
25 kDa (SNAP-25), and synaptotagmin 1 after an insulin secretiontest under exposure to an ELFMF or under sham conditions. Twofold serial dilutions of cDNA were amplified for 25 (insulin 1),
(bp), respectively), and PCR products were undetect-
29 (insulin 2 and SNAP-25), 33 (synaptotagmin 1) or 26 (b-actin)
able when reverse transcription was performed without
cycles. Lanes 1^ 3 represent serial dilutions of cDNA sample.
reverse transcriptase (ThermoScript RT). Semi-quanti-
Lane1was the most concentrated sample in each series. B: Semi-
tative RT-PCR was performed under conditions where
quantitative RT-PCR analysis of the effects of exposure to anELFMF on expression of mRNA encoding insulin 1, insulin 2,
the amplification reaction for the PCR products was
SNAP-25, and synaptotagmin 1. Insulin secretion tests were per-
within the linear range. For example, the increase in
formed using stimulation buffer containing 3.3 mM D-glucose or
the optical density of the amplified PCR products for
3.3 mM D-glucose and 45 mM KCl. Data represent the mean Æ SE
b-actin was linear between at least cycles 24 and 29. A
(insulin 1 and insulin 2, n ¼ 3; SNAP-25, n ¼ 5; synaptotagmin 1,
reduced intensity of products was observed at each
dilution step (Fig. 4A). This result confirmed that theRT-PCR was performed in the exponential portion of
In the presence of 45 mM KCl, exposure to an
Recently, human exposure to ELFMFs from vari-
ELFMF significantly reduced expression of the mRNA
ous electrical appliances has increased significantly.
encoding SNAP-25 and synaptotagmin 1 by approxi-
Thus, the possible health effects of exposure to ELFMF
mately 36 and 23%, respectively, compared to sham
have become a considerable public concern. On the
exposed cells. Insulin 2 mRNA expression was slightly,
other hand, occurrence of diabetes mellitus has increas-
but not significantly, reduced by exposure to ELFMF
ed progressively in recent years. Insufficient pancreatic
(approximately 10%), and insulin 1 mRNA expression
islet function is the basis of various forms of diabetes.
did not decrease. Insulin 2, SNAP-25, and synaptotag-
These two factors motivated us to investigate the effects
min 1 mRNA expression increased when RIN-m cells
of exposure to ELFMF on the function of insulin
were stimulated by 45 mM KCl, compared to 3.3 mM
secreting cells. If exposure to ELFMF is deleterious
to insulin secreting cells, it is essential that this is
demonstrated and communicated as soon as possible. In
brane. SNARE proteins play a role in this tethering/
contrast, if exposure to ELFMF is beneficial to insulin
docking process between secretory granules and the
secreting cells, it might be possible to utilize exposure
plasma membrane. SNAP-25 is a SNARE protein that
to ELFMF for medical applications. For example, an
is expressed in pancreatic islets and is involved in in-
ELFMF might be used in diabetes mellitus to decrease
sulin release [Sadoul et al., 1995; Wheeler et al., 1996].
blood glucose levels, and to increase insulin levels
SNAP-25 mRNA expression was significantly increas-
in blood [Laitl-Kobierska et al., 2002]. Because of these
ed when chromaffin cells [Garcia-Palomero et al., 2000;
issues, the assessment of the effects of exposure to
Montiel et al., 2003] and rat granulosa cells [Grosse
ELFMF on insulin secretion is very important.
et al., 2000] were stimulated to release neurotransmit-
Studies evaluating the influence of exposure to
ters, and these events were closely related to the influx
ELFMF on pancreatic islet function are scarce, and it is
often difficult to compare the studies that have been
Synaptotagmin 1 is a calcium ion sensor protein
performed because of the different research methods
that is located on the membrane of insulin containing
used. Hence, an association between magnetic field
secretory granules [Lang et al., 1997]. It is thought to be
exposure and pancreatic islet function has not been
play a role in the fusion between secretory granules and
demonstrated unequivocally. Furthermore, the use of
the plasma membrane according to the elevation of the
islets results in significant methodological difficulties,
cellular calcium concentration [Gerber and Sudhof,
due to cellular heterogeneity, limited availability, and
rapid deterioration of function. In order to circumvent
In this work, the expression of mRNA encoding
these problems in the current study, we used insulinoma
SNAP-25 and synaptotagmin 1 decreased under ex-
cells instead of islets. To our knowledge, this is the first
posure to an ELFMF in the presence of 45 mM KCl.
investigation in which insulinoma cells have been used
From these findings, we conclude that exposure to
to examine the effects of exposure to ELFMF on insulin
ELFMF attenuates insulin secretion by reducing the
secreting cells. The advantages of using insulinoma
influx of calcium ions through calcium channels. It has
cells were the ease of generation of large quantities of
been reported that calcium ion content, calcium ion
functional cells, and the stability of the resulting cell
efflux, and insulin secretion during glucose stimulation
was reduced when isolated rabbit islets were exposed to
We investigated the insulinoma cell line, RIN-m.
low frequency pulsed magnetic fields [Joelly et al.,
It has been reported that PMA, which acts via the PKC
1983]. It has also been reported that exposure to
pathway, induces insulin secretion [Yada et al., 1989].
ELFMF affected neurite growth via voltage gated
On the basis of the insulin secretory response of RIN-m
calcium channels [Morgado-Valle et al., 1998] and af-
cells to 45 mM KCl and PMA, we thought that RIN-m
fected the differentiation of neuroblastoma cells by
cells were suitable for a model to evaluate the effects of
antagonizing the shift in cell membrane surface charges
exposure to ELFMF on the potassium induced insulin
and increasing intracellular calcium levels [Tonini et al.,
secretion pathway and the PKC cascade. A high con-
2001]. Our present findings are in agreement with these
centration of D-glucose induces the closure of ATP de-
pendent potassium channels in insulin secreting cells,
In the current study, exposure to an ELFMF
which in turn induces membrane depolarization, open-
slightly increased PMA stimulated insulin secretion,
ing of voltage dependent calcium channels, and insulin
but no statistically significant difference was observed
secretion. It has been reported that treatment with high
at 5 mT and 60 Hz. We have previously reported that
concentrations of potassium bypasses the ATP depen-
exposure to ELFMF at 50 Hz and 400 mT, but not
dent potassium channels of insulin secreting cells and
at 5 mT, enhanced the expression of a neuron deriv-
also induces insulin secretion [Hohmeier et al., 2000].
ed orphan receptor gene induced by treatment with
In the current study, insulin secretion induced by
forskolin and PMA [Miyakoshi et al., 1998]. We also
45 mM KCl was attenuated by approximately 30%
reported that the suppression of heat shock protein 70
under exposure to an ELFMF, compared to sham ex-
was observed at a magnetic density of 50 mT, but not at
posure. Treatment with nifedipine reduced the differ-
5 or 0.5 mT [Miyakoshi et al., 2000]. These previous
ence in insulin secretion between ELFMF exposed and
results indicate the dependence of the effects of ex-
sham exposed cells. Recent investigations have clarifi-
posure to ELFMF on the strength of the magnetic field,
ed the molecular mechanisms of exocytosis in neuro-
and they are consistent with the results of the current
transmitter release or hormone secretion [Jones and
Persaud, 1998; Gerber and Sudhof, 2002]. In the exo-
The results presented here suggest that insulin
cytotic release process, secretory granules are docked at
secretion decreases under exposure to ELFMF. Hence,
the site of exocytosis and fused with the plasma mem-
it might be desirable for diabetic patients who have
insufficient insulin secretion from pancreatic islets to
Joelly WB, Hinshaw DB, Knierim K. 1983. Magnetic field effects
on calcium efflux and insulin secretion in isolated rabbit
In summary, we investigated the effects of ex-
islets of Langerhans. Bioelectromagnetics 4:103–106.
Jones PE, Persaud SJ. 1998. Protein kinase, protein phosphoryla-
posure to ELFMF on insulin secretion using the in-
tion, and the regulation of insulin secretion from pancreatic
sulinoma cell line, RIN-m. In the presence of 45 mM
KCl, exposure to ELFMF significantly attenuated in-
Laitl-Kobierska A, Cieslar G, Sieron A, et al. 2002. Influence of
sulin release from RIN-m cells, compared to sham
alternating extremely low frequency ELF magnetic field on
exposed cells. Treatment with nifedipine reduced the
structure and function of pancreas in rats. Bioelectromag-netics 23:49–58.
difference in insulin secretion between ELFMF ex-
Lang J, Fukuda M, Zhang H, et al. 1997. The first C2 domain of
posed and sham exposed cells, and the expression of
synaptotagmin is required for exocytosis of insulin from
mRNA encoding SNAP-25 and synaptotagmin 1 de-
pancreatic b-cells: Action of synaptotagmin at low micro-
creased under exposure to an ELFMF in the presence of
molar calcium. EMBO J 16:5837–5846.
45 mM KCl. SNAP-25 is important in tethering and
Miyakoshi J, Ohtsu S, Shibata T, et al. 1996. Exposure to magnetic
field (5 mT at 60 Hz) does not affect cell growth and c-myc
docking between secretory granules and plasma mem-
gene expression. J Radiat Res 37:185–191.
branes and these events are closely related to the influx
Miyakoshi J, Tsukada T, Tachiiri S, et al. 1998. Enhanced NOR-1
of calcium ions. Synaptotagmin 1 is a calcium ion
gene expression by exposure of Chinese hamster cells to
sensor protein. These results suggest that exposure to
high-density 50 Hz magnetic fields. Mol Cell Biochem 181:
ELFMF attenuates insulin secretion from RIN-m cells
Miyakoshi J, Mori Y, Yaguchi H, et al. 2000. Suppression of heat-
by affecting calcium influx through calcium channels.
induced HSP-70 by simultaneous exposure to 50 mT magne-tic field. Life Sci 66:1187–1196.
Montiel C, Mendoza I, Garcia CJ, et al. 2003. Distinct protein kinase
regulate SNAP-25 expression in chromaffin cells. J Neurosci
Bhathena SJ, Awoke S, Voyles NR, et al. 1984. Insulin, glucagon,
and somatostatin secretion by cultured rat islet cell tumor and
Morgado-Valle C, Verdugo-Diaz L, Garcia DE, et al. 1998. The
its clones (41762). Proc Soc Exp Biol Med 175:35–38.
role of voltage-gated Ca2þ channels in neurite growth of
Chick WL, Warren S, Chute RN, et al. 1977. A transplantable
cultured chromaffin cells induced by extremely low fre-
insulinoma in the rat. Proc Natl Acad Sci 74:628–632.
quency (ELF) magnetic field stimulation. Cell Tissue Res
Ding D-R, Yaguchi H, Yoshida M, et al. 2000. Increase in X-ray-
induced mutations by exposure to magnetic field (60 Hz,
Praz GA, Halban PA, Wollheim CB, et al. 1983. Regulation of im-
5 mT) in NF-kB-inhibited cells. Biochem Biophys Res
munoreactive-insulin release from a rat cell line (RINm5F).
Garcia-Palomero E, Montiel C, Herrero CJ, et al. 2000. Multiple
Sadoul K, Lang J, Montecucco C, et al. 1995. SNAP-25 is expressed
calcium pathways induce the expression of SNAP-25 protein
in islets of Langerhans and is involved in insulin release.
in chromaffin cells. J Neurochem 74:1049–1058.
Gazdar AF, Chick WL, Oie HK, et al. 1980. Continuous, clonal,
Savitz DA, Loomis DP. 1995. Magnetic field exposure in relation to
insulin- and somatostatin-secreting cell lines established
leukemia and brain cancer mortality among electric utility
from a transplantable rat islet cell tumor. Proc Natl Acad Sci
workers. Am J Epidemiol 141:123–134.
Tonini R, Baroni MD, Masala E, et al. 2001. Calcium protects
Gerber SH, Sudhof TC. 2002. Molecular determinants of regulated
differentiating neuroblastoma cells during 50 Hz electro-
magnetic radiation. Biophys J 81:2580–2589.
Grosse J, Bulling A, Brucker C, et al. 2000. Synaptosome-
Wheeler MB, Sheu L, Ghai M, et al. 1996. Characterization of
associated protein of 25 kilodaltons in oocytes and steroid-
SNARE protein Expression in b-cell lines and pancreatic
producing cells of rat and human ovary: Molecular analysis
islets. Endocrinology 137:1340–1348.
and regulation by gonadotropins. Biol Reprod 63:643–650.
Yada T, Russo LL, Sharp WG. 1989. Phorbol ester-stimulated
Hayek A, Guardian C, Guardian J, et al. 1984. Homogeneous
insulin secretion by RINm5F insulinoma cells is linked with
magnetic fields influence pancreatic islet function in vitro.
membrane depolarization and an increase in cytosolic free
Biochem Biophys Res Commun 122:191–196.
Ca2þ concentration. J Biol Chem 264:2455–2462.
Hohmeier HE, Mulder H, Chen G, et al. 2000. Isolation of INS-1-
Yaguchi H, Yoshida M, Ding D-R, et al. 2000. Increased chromatid-
derived cell lines with robust ATP-sensitive Kþ channel-
type chromosomal aberrations in mouse m5S cells exposed to
dependent and -independent glucose-stimulated insulin
power-line frequency magnetic fields. Int J Radiat Biol 76:
Slashdot | Social Consequences and Effects of RFID Implants?http://ask.slashdot.org/article.pl?sid=06/05/04/0030212 Slashdot News for nerds, stuff that matters Social Consequences and Effects of RFID Implants? Posted by Cliff on Wednesday May 03, @11:45PM from the chips-in-my-head dept. kramdam asks: "Even with all the talk about privacy and security, there seems to be a growi
Washington D.C. -Mandatory Meeting March 27 6:30 p.m. o Sign parent and student name on sign in sheeto Pick up the agenda/information packet, room & bus lists, itinerary and 2 luggage tagso Chaperones leave for chaperone meeting w/Mrs. Johnson – room 123o Picasa Photos - Pathfinder homepage, Activities then DC trip. o Twitter – Pathfinder homepage, Activities then DC Tripo Group
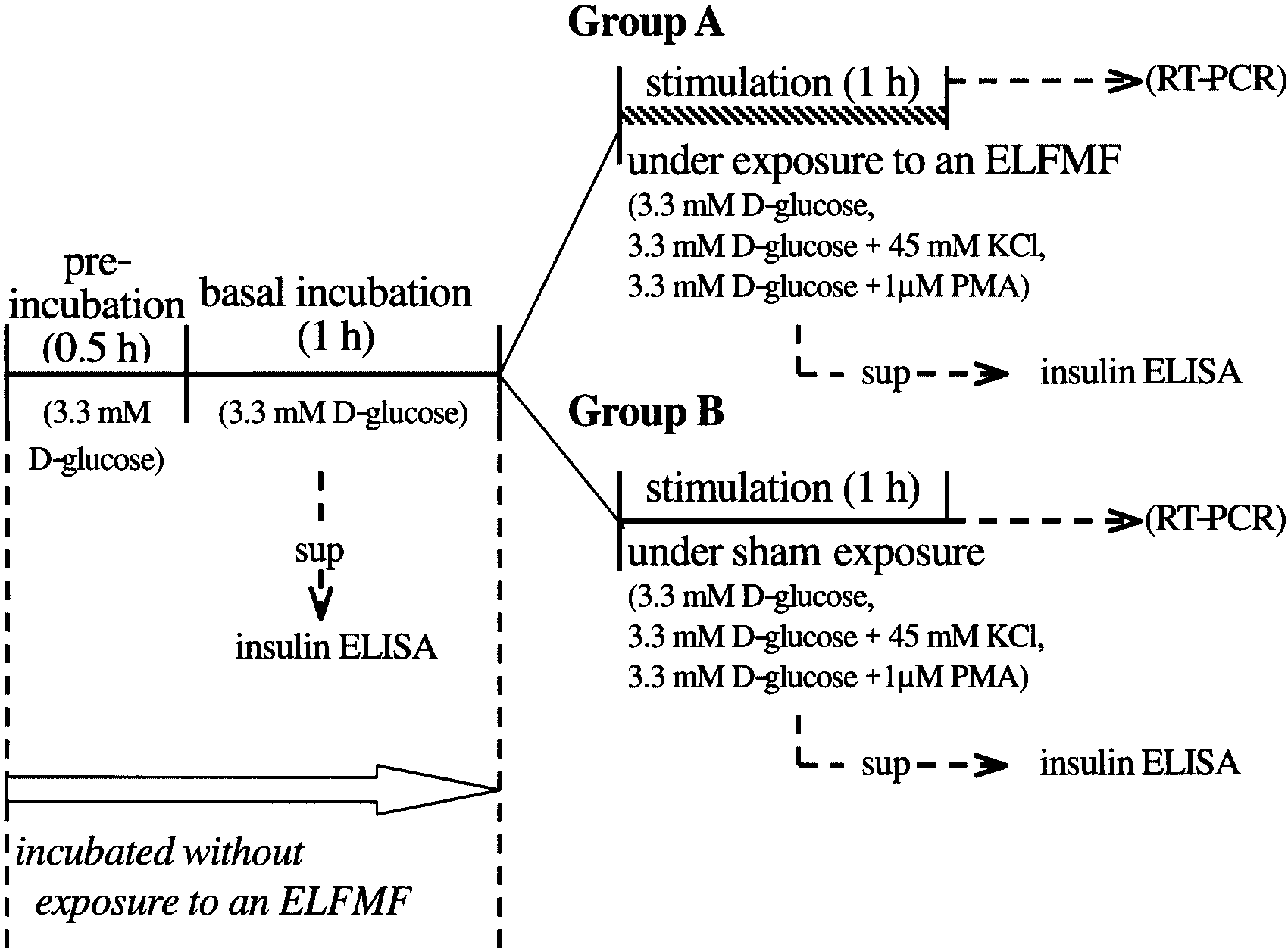 Fig. 1. The outline of insulin secretion tests. Preincubation and basal incubation were performedwithoutexposure to anextremelylow frequencymagnetic field (ELFMF).After wellswere dividedintotwo groups, one group is stimulated under exposure to an ELFMF (Group A), and the other group isstimulatedunder sham exposure (Group B). At the end ofthe basalincubation period and stimulationperiod, an aliquot of the incubation buffer in each well was collected, and the insulin concentration ofthe samples was measured by enzyme linked immunosorbent assay (ELISA).
Fig. 1. The outline of insulin secretion tests. Preincubation and basal incubation were performedwithoutexposure to anextremelylow frequencymagnetic field (ELFMF).After wellswere dividedintotwo groups, one group is stimulated under exposure to an ELFMF (Group A), and the other group isstimulatedunder sham exposure (Group B). At the end ofthe basalincubation period and stimulationperiod, an aliquot of the incubation buffer in each well was collected, and the insulin concentration ofthe samples was measured by enzyme linked immunosorbent assay (ELISA).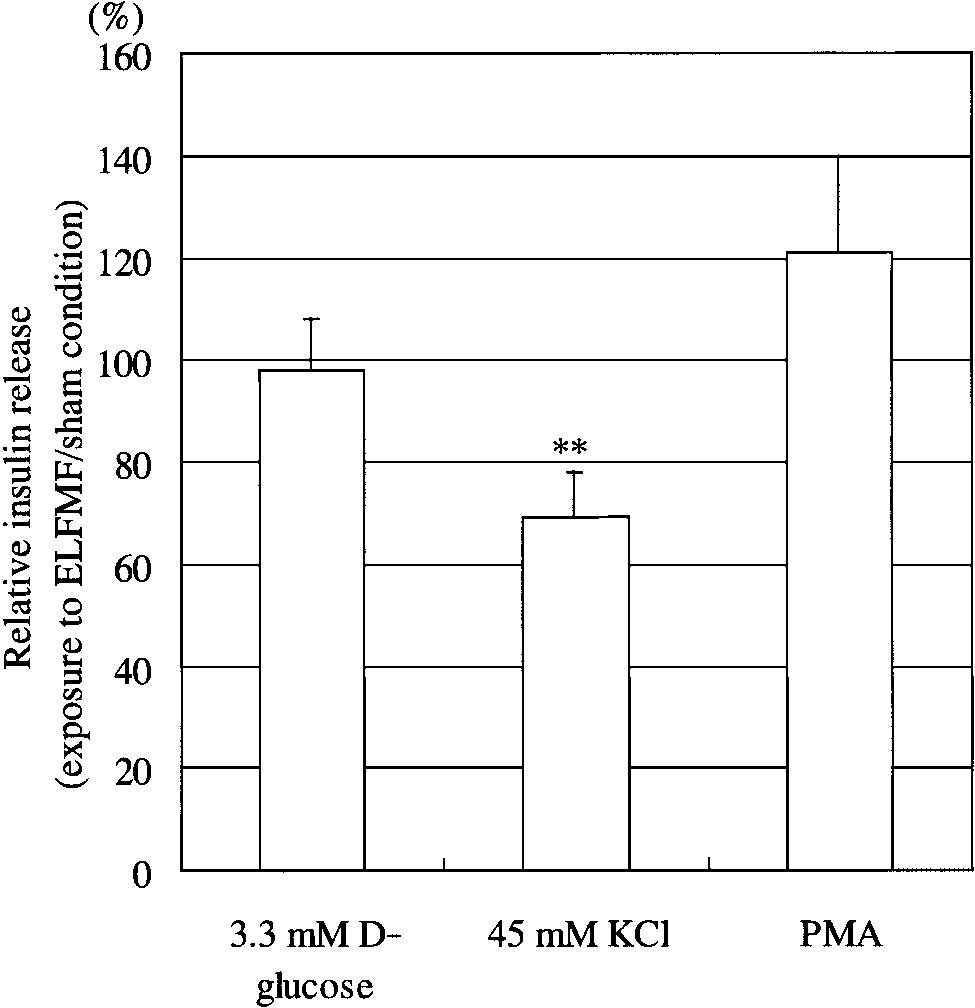 Insulin Secretion From RIN-m CellsUnder Sham Conditions
Under sham conditions, incubation with stimula-
tion buffer containing a high concentration of potas-sium (45 mM KCl) caused insulin release to increaseapproximately 14 fold, compared to the basal incuba-tion conditions. In contrast, both a normal (3.3 mM)and a high (16.7 mM) concentration of D-glucose didnot increase insulin release, compared to the basal in-cubation conditions. Phorbol-12-myristate-13-acetate(PMA) also increased insulin release from RIN-m cellsby approximately 15 fold (Table 1). These results arein agreement with previous reports describing RIN-mcells and their subcloned cell line, RINm5F [Praz et al.,1983; Bhathena et al., 1984; Yada et al., 1989].
Insulin Secretion From RIN-m CellsUnder Sham Conditions
Under sham conditions, incubation with stimula-
tion buffer containing a high concentration of potas-sium (45 mM KCl) caused insulin release to increaseapproximately 14 fold, compared to the basal incuba-tion conditions. In contrast, both a normal (3.3 mM)and a high (16.7 mM) concentration of D-glucose didnot increase insulin release, compared to the basal in-cubation conditions. Phorbol-12-myristate-13-acetate(PMA) also increased insulin release from RIN-m cellsby approximately 15 fold (Table 1). These results arein agreement with previous reports describing RIN-mcells and their subcloned cell line, RINm5F [Praz et al.,1983; Bhathena et al., 1984; Yada et al., 1989].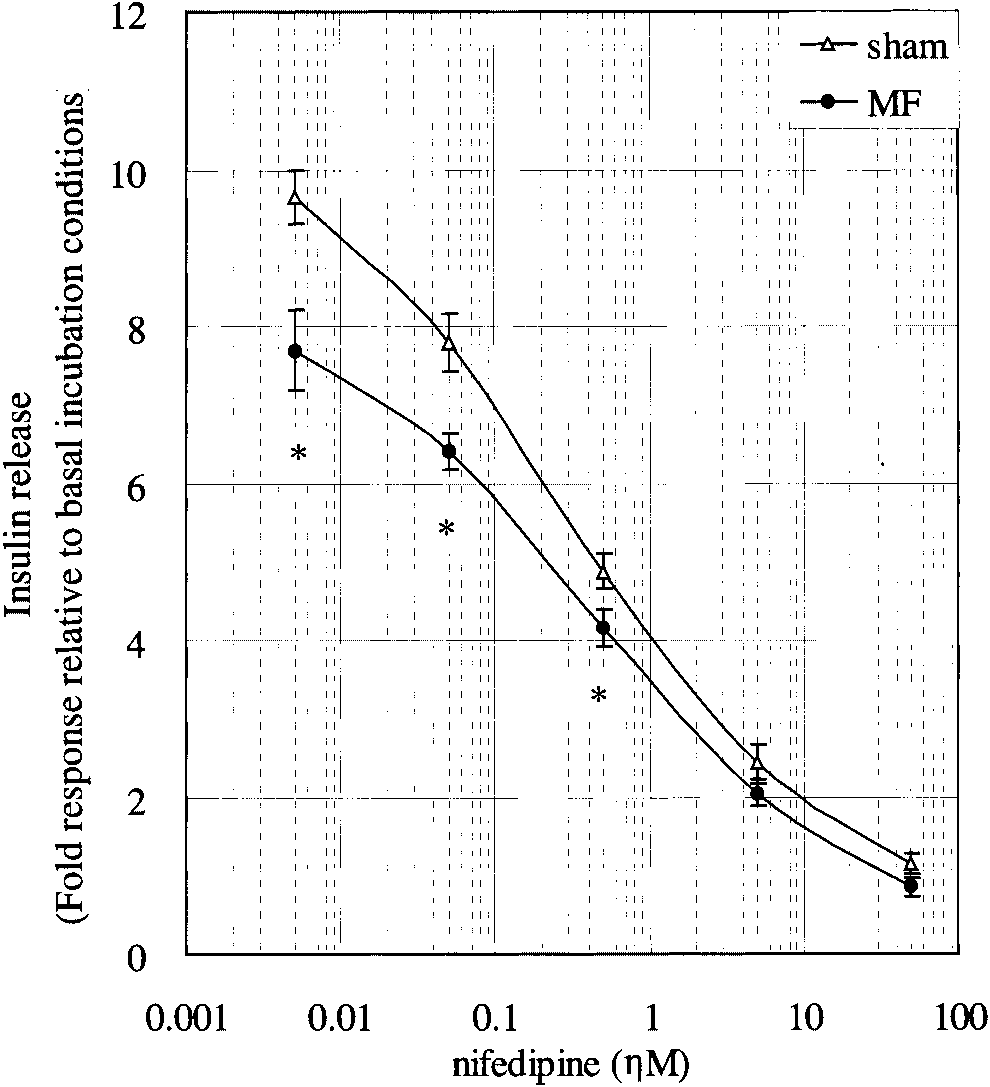
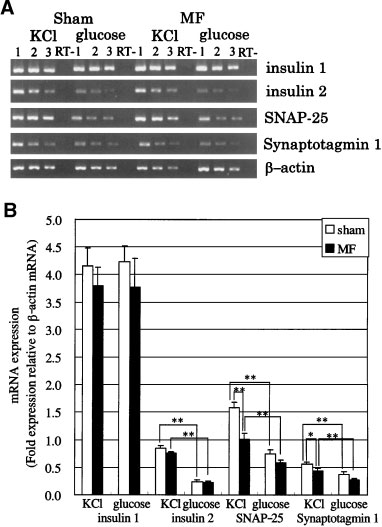 Fig. 3. Nifedipine inhibition of insulin release from RIN-m cells sti-mulated by 3.3 mM D-glucose and 45 mMKClunderexposure to anELFMF or under sham conditions.To study the effect of nifedipine,cells were incubated with the indicated concentrations of nifedi-pine for 30 min, and then incubated for 1 h with stimulation buffercontaining 3.3 mM D-glucose and 45 mM KCl. Each point repre-
Fig. 4. A: Reverse transcription-polymerase chain reaction (RT-
sents the mean Æ SE (50 mM nifedipine, n ¼ 3; other concentra-
PCR) of insulin 1, insulin 2, synaptosomal associated protein of
25 kDa (SNAP-25), and synaptotagmin 1 after an insulin secretiontest under exposure to an ELFMF or under sham conditions.
Fig. 3. Nifedipine inhibition of insulin release from RIN-m cells sti-mulated by 3.3 mM D-glucose and 45 mMKClunderexposure to anELFMF or under sham conditions.To study the effect of nifedipine,cells were incubated with the indicated concentrations of nifedi-pine for 30 min, and then incubated for 1 h with stimulation buffercontaining 3.3 mM D-glucose and 45 mM KCl. Each point repre-
Fig. 4. A: Reverse transcription-polymerase chain reaction (RT-
sents the mean Æ SE (50 mM nifedipine, n ¼ 3; other concentra-
PCR) of insulin 1, insulin 2, synaptosomal associated protein of
25 kDa (SNAP-25), and synaptotagmin 1 after an insulin secretiontest under exposure to an ELFMF or under sham conditions.