Tadalafil appartiene alla classe degli inibitori selettivi della fosfodiesterasi di tipo 5, con un profilo farmacocinetico caratterizzato da un’emivita terminale di circa diciotto ore. Dopo somministrazione orale viene assorbito rapidamente e raggiunge concentrazioni plasmatiche massime in due ore. La biotrasformazione avviene principalmente tramite CYP3A4 con formazione di metaboliti inattivi, escreti in prevalenza con le feci. L’elevato legame alle proteine plasmatiche (>90%) assicura una distribuzione stabile. Nei confronti delle altre molecole della stessa classe, cialis compresse italia è noto per la durata prolungata dell’attività farmacologica.
Tavernarakislab.gr
Insect Molecular Biology (2006) 15(1), 95–103 Germ line transformation of the olive fly Bactrocera oleae using a versatile transgenesis marker M. Koukidou1*†, A. Klinakis1*†‡, C. Reboulakis§,
presents an increasing challenge to pest control. Progress
L. Zagoraiou*†¶, N. Tavernarakis*, I. Livadaras*,
in molecular manipulation of insect species is an important
A. Economopoulos§ and C. Savakis*†
way to further comprehension of the genetic and biochemical
*Institute of Molecular Biology and Biotechnology,
basis of insect biology. In addition, it will open the way for
Foundation for Research and Technology, Heraklion,
novel strategies to control insect pest populations, as, for
Crete, Greece; †Medical School and §Department of
example, through production of insects for sterile insect
Biology, University of Crete, Heraklion, Greece
The technologies for insect transgenesis developed so
far rely on transposable elements. Manipulation of an insect
Abstract
genome with a transposable element was first achieved in
The olive fruit fly (olive fly) Bactrocera oleae (Dacus), Drosophila with the P element (Rubin & Spradling, 1982). recently introduced in North America, is the most
However, the host range of P seems to be restricted to the
destructive pest of olives worldwide. The lack of an
drosophilids (Handler et al., 1993). The discovery of new
efficient gene transfer technology for olive fly has
elements with a broader host range has led to the develop-
hampered molecular analysis, as well as development
ment of vector systems for transformation of several non-
of genetic techniques for its control. We have developed
drosophilid insect species, including the Mediterranean fruit
a Minos-based transposon vector carrying a self-
fly Ceratitis capitata (Loukeris et al., 1995b; Handler et al.,
activating cassette which overexpresses the enhanced
1998; Michel et al., 2001), the mosquitoes Aedes aegyptigreen fluorescent protein (EGFP). Efficient transposase-
(Jasinskiene et al., 1998; Coates et al., 1998), Anophelesmediated integration of one to multiple copies of this albimanus (Perera et al., 2002), Anopheles stephensivector was achieved in the germ line of B. oleae by
(Catteruccia et al., 2000) and Anopheles gambiae (Grossman
coinjecting the vector along with in vitro synthesized et al., 2001), the housefly Musca domestica (Hediger et al.,
Minos transposase mRNA into preblastoderm embryos.
2001), the Australian sheep blowfly Lucilia cuprina (Heinrich
The self-activating gene construct combined with trans- et al., 2002), the silkmoth Bombyx mori (Tamura et al.,
posase mRNA present a system with potential for
2000), the beetle Tribolium castaneum (Lorenzen et al.,
transgenesis of very diverse species.
2003; Pavlopoulos et al., 2004) and the New World screw-worm Cochliomyia hominivorax (Allen et al., 2004). Keywords: Olive fruit fly; Bactrocera oleae; germ line Bactrocera oleae is the main pest of olives in the
transformation; transgenesis marker.
Mediterranean region and in California, where it has beenintroduced recently. Currently, control of B. oleae is based
Introduction
on the use of insecticides either in bait or in cover sprays,resulting in adverse effects on the environment and some-
Arthropods are key pests of food and fibre crops as well
times presence of unacceptably high levels of insecticides
as major disease vectors. Field resistance to pesticides
in olives and olive oil (Ferreira & Tainha, 1983). Develop-ment of olive fly control strategies leading to reduced or no
use of insecticides has been attempted with mixed results,
Received 18 April 2005; accepted after revision 29 August 2005. Corre-spondence: Charalambos Savakis, Medical School, University of Crete
for example, ‘lure and kill’ (Haniotakis et al., 1991), localized
and IMBB-FoRTH, PO Box 1527, Heraklion 71110, Crete, Greece.
bait spray (Pucci, 1990) and inundative or inoculative
Tel.: +30 2810391114, fax: +30 2810391950, E-mail: [email protected]
release of parasitoids (Tzanakakis, 1995). SIT has been
Current addresses: ‡Department of Genetics and Development, Columbia
used with limited success (Economopoulos et al., 1977),
University, NY, USA, ¶Center for Neurobiology and Behaviour, ColumbiaUniversity, NY, USA
apparently because of low competitiveness of the mass-
1These authors contributed equally to this work.
reared males compared to the wild males (Economopoulos
& Zervas, 1982). Availability of a method for olive flytransgenesis is a prerequisite for genetic manipulation ofthis species for development of more effective controlmethods, such as release of insects carrying a dominantlethal allele (RIDL) strategies (Heinrich & Scott, 2000;Thomas et al., 2000) and sensitization of insect populationsto pro-insecticides (SIPP) (Markaki et al., 2004).
The transposon Minos from Drosophila hydei belongs
to the Tc1/Mariner family of transposable elements (Franz& Savakis, 1991) which, in addition to insects, has beenshown to mediate transgenesis in human cells (Klinakiset al., 2000a), in mouse somatic and germ cells (Zagoraiouet al., 2001; Drabek et al., 2003) and in the ascidian Cionaintestinalis (Sasakura et al., 2003). Encouraged by thisbroad host range, we attempted germ-line transformationof B. oleae with a Minos vector carrying an enhanced greenfluorescent protein (EGFP) gene. EGFP is a universalmarker for insect transgenesis (Berghammer et al., 1999).
In this report, we describe a self-stimulating transcription
system for high levels of regulatable expression of markergenes in diverse species. Using this system in combinationwith a Minos transposon, we demonstrate stable and efficienttransgenesis of B. oleae. A potentially universal EGFP-based genetic marker
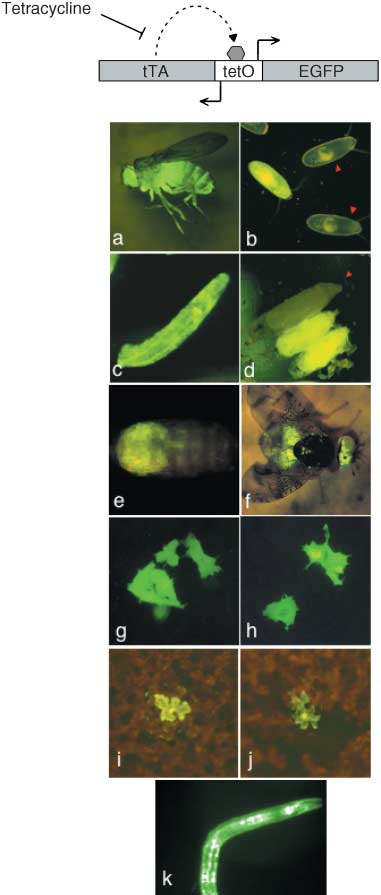
To achieve high levels of EGFP expression, we used aself-activating and self-sustained positive feedback loopbased on the tetracycline-sensitive transcriptional activatortTA (Gossen & Bujard, 1992), which is based on con-comitant expression of EGFP and tTA from a bi-directionalpromoter driven by tTA. Two versions of the expressioncassette were generated. The first version was derivedfrom a commercially available cassette consisting of a 7-merof the tetracycline operator (tetO) (Gossen & Bujard, 1992)flanked by two divergent copies of a cytomegalovirus(CMV) minimal promoter (the ‘mammalian version’). Thesecond was constructed for use in insects and consistedof a 14-mer of the tetO element flanked by two divergentcopies of the minimal promoter of the Drosophila mela-nogaster hsp70 gene (the ‘insect version’). A 14-mer wasused in the insect version in order to maximize expressionlevels (Fig. 1, top).
The insect cassette was tested in D. melanogaster and in
the medfly Ceratitis capitata by P- and Minos-based germline transformation, respectively. In D. melanogaster, most
Figure 1. Top: the tTA/enhanced green fluorescent protein (EGFP) self-stimulating expression cassette. Bottom: EGFP expression.
of the transgenic lines exhibited strong, ubiquitous and
(a–d) transformed Drosophila melanogaster adults, embryos, third instar
constitutive expression of EGFP throughout development
larva and pupae; red arrowheads point to non-transformed individuals.
(Fig. 1a – d), while only some lines showed tissue-specific
(e–f) Transformed medfly pupa and adult. (g–h) Human HeLa cells stably transfected with insect and mammalian cassette, respectively. (i–j) Nicotiana
expression patterns (not shown). Fluorescence was less
tabacum leaf transiently transfected with insect and mammalian cassette,
intense in medfly transformants, which were characterized
respectively. (k) Transgenic Caenorhabditis elegans animal harbouring a
by specific patterns of expression during development
plasmid vector carrying the insect cassette.
2006 The Royal Entomological Society, Insect Molecular Biology, 15, 95–103 Germ line transformation of the olive fruit fly
The versatility of the bidirectional loop was further
explored in somatic cells of animals and plants. HeLa cells
Olive fly transformation with a Minos element
were transiently and stably transfected with both the
Preblastoderm embryos of the B. oleae Demokritos strain
‘mammalian’ and the ‘insect’ construct; fluorescence was
were coinjected with the Minos-based ‘insect’ construct and
detectable 24 h post transfection (Fig. 1g–h) and peaked
Minos transposase mRNA, a method that has been previ-
48 h post-transfection. The same constructs were also
ously shown to result in high transformation frequencies
introduced into tobacco leaves (Nicotiana tabacum and
(Kapetanaki et al., 2002). Transient EGFP expression was
N. bentamiana) via gold particle ‘bombardment’; examination
observed in about 60% of the injected embryos examined
48 h post transfection showed strong EGFP expression in
(Fig. 2, panels A to C), indicating that the genetic marker is
certain cells (Fig. 1i – j). The ‘insect’ vector was also intro-
duced into the nematode Caenorhabditis elegans, leading
From the 3833 injected G0 embryos, 641 larvae hatched,
to strong and uniform EGFP expression (Fig. 1k).
from which 151 male and 174 female flies developed. The
Experiments performed in Drosophila and in HeLa cells
male G0 flies were mated in a single cage to 506 non-
carrying stable, active integrations of the tTA-containing
injected female flies, and the female G0 flies were mated in
cassette showed that EGFP expression is reversibly sup-
another cage to 250 wild-type male flies. The G1 progeny
pressed by the teracycline analogue doxycycline at con-
from the two crosses was screened for EGFP fluorescence
centrations known to inhibit tTA binding to the promoter
at the third instar larval stage. A total of 8824 third instar
(data not shown). Furthermore, high levels of EGFP expres-
larvae was screened; 67 of them exhibited green fluores-
sion were dependent on the presence of the bidirectional
cence in various tissues and were therefore considered
loop because no fluorescence was obtained from control
constructs containing the bidirectional promoter without the
To establish transformed lines, 13 individual G1 flies
were back-crossed to wild-type flies and the resulting G2
Figure 2. Patterns of enhanced green fluorescent protein (EGFP) expression in injected embryos and in transformed Bactrocera oleae individuals. (a–c) Injected eggs showing various patterns of transient EGFP expression. (d–f) EGFP expression in larva, pupa and adult G2 progeny from line 12. (g–i) EGFP expression in larva, pupa and adult G2 progeny from line 10. ( j–l) EGFP expression in larva, pupa and adult G2 progeny from line 9. (m–o) Nontransformed larva, pupa and adult.
2006 The Royal Entomological Society, Insect Molecular Biology, 15, 95–103 Figure 3. Frequencies of transformants among the olive fly G2 progeny. enhanced green fluorescent protein (EGFP)-expressing G1 progeny was individually backcrossed to wild-type flies. The G2 progeny was screened for EGFP expression. Bars indicate the total number of G2 flies Figure 4. Southern blot analysis of transformed olive fly lines. Top:
from each G1 parent. The proportion of G2 progeny expressing the EGFP
schematic of the ‘insect version’ transposon. The bar indicates the fragment
marker is also indicated. No G2 progeny were obtained from cage numbers
used as probe for Southern blot analysis. Bottom: Southern blots of the
EcoRI digested genomic DNA from individual enhanced green fluorescent protein (EGFP)-expressing G2 progeny. Line numbers are indicated at the top of the panel. Marker values are in kilobases.
third instar larvae were again screened for EGFP expression. Four of the crosses did not give any progeny. Where numbers
Molecular analysis of the transformed lines
allowed assessment, the proportion of progeny expressing
Insertion of the Minos element into the B. oleae genome
the EGFP marker was consistent with segregation of a
was verified by Southern blot analysis on EcoRI-digested
genomic DNA with an EGFP probe. Any single, transposase-
Clearly detectable EGFP fluorescence was observed in
dependent insertion of the transposon into genomic DNA is
second and third instar larvae, pupae and adult flies in all
expected to give a band of at least 1300 bp in size. Based
transformed lines examined. Nine independent transformed
on this criterion, the presence of a single insertion was
lines were established (lines 2, 5, 6, 8, 9, 10, 11, 12 and 13).
detected in progeny of transgenic flies 2, 5 and 6, while
The pattern of EGFP expression differed considerably from
progeny of transgenics 8, 9, 10, 11, 12 and 13 contained
line to line (Fig. 2). Furthermore, some lines showed vari-
multiple insertions (Fig. 4). Of the nine cases analysed,
ability of EGFP expression (variegation) between siblings,
only two, 5 and 6, were found to yield a band of the same
although the overall pattern was characteristic (lines 9, 11
size (Fig. 4). Additionally, these two showed the same pat-
and 13; data not shown). With one exception (line 9),
tern of EGFP expression, suggesting that they represent
patterns of expression did not change through the genera-
the same transformation event. The others were regarded
tions. At least two different expression patterns were
as independent transformation events. Southern blot ana-
detected and were segregating in line 9, one of which,
lysis performed in individual flies from lines 13 and 11 at
highly localized, is shown in Fig. 2(j – l). Because line 9
subsequent generations showed that some of the bands
contained multiple insertions, it is likely that two active
were lost, clearly suggesting that individual bands repre-
insertions (i.e. insertions expressing EGFP) existed in this
sent independent, unlinked integrations of the element
line, each with a distinct pattern of expression. Transmis-
sion of the EGFP marker has been maintained in all lines
To verify that the insertions are indeed transposase-
for at least eight generations, demonstrating that the
mediated, integration sites were amplified by inverse PCR
marker is stably inherited, as expected in the absence of
(Triglia et al., 1988) from four of the lines. The PCR prod-
ucts were cloned and sequenced. As shown in Fig. 5, each
Binding of the transcriptional activator tTA to its target is
of the four integration sites contains the Minos inverted ter-
inhibited by tetracycline (Gossen & Bujard, 1992). Tetracycline-
minal repeat followed by a TA dinucleotide and a sequence
dependence of EGFP expression was tested in one of the
not present in the original plasmid. We therefore conclude
lines (line 12), which is characterized by high levels of
that the Minos construct inserts through transposition and
fluorescence in all stages. Fluorescence is not detectable
in larvae of line 12 grown on medium containing 10 mg / l
The sequences adjacent to the insertion sites were
subjected to BLAST analysis against GENBANK. In two of the
2006 The Royal Entomological Society, Insect Molecular Biology, 15, 95–103 Germ line transformation of the olive fruit flyFigure 5. Sequences flanking Minos insertions. The end of the transposon sequence is in bold face. The MboI restriction site used in inverse PCR is underlined. The TA target dinucleotide is indicated.
sites analysed, the retrieved flanking sequences were too
for transformants, which was performed in the absence of
short to allow a meaningful BLAST analysis (Fig. 5). One of
tetracycline and was based on EGFP expression. We
the two other sequences (line 10) showed a 91% identity
observed, nevertheless, tetracycline-repressible lethality
(E-value: 2e-11) to a region within the large intron of the
in a number of Drosophila transformants, only in the
corkscrew gene of Drosophila (see Supplementary Material).
homozygous form. As we did not perform a systematicscreen for dominant tet-repressible lethals, we cannot estim-ate the frequency of tTA-induced lethal transgenes carrying
Discussion
our bidirectional tTA/EGFP construct. It is also possible that
We have constructed a versatile marker cassette, in which
the strong lethality observed by Gong et al. (2005) may be
the EGFP gene is expressed from a bidirectional promoter
caused by higher levels of tTA, as a tTA variant was used
controlled by the synthetic transcriptional activator tTA
which is optimized for expression in D. melanogaster.
(Gossen & Bujard, 1992); the promoter also drives the
Using the tTA/EGFP self-sustaining cassette, we have
expression of tTA. This results in self-sustaining expression
shown that the transposable element Minos from D hydei
of the tTA activator. This positive-feedback system should
can mediate stable germ-line transformation of B. oleae,
be constitutively active once transcription is initiated, for
thus extending the number of species that have been
example from an enhancer near the site of insertion.
transformed with this element. Transformation frequency,
The marker cassette presents several interesting fea-
expressed as the fraction of G0 individuals producing trans-
tures. Firstly, strong EGFP expression in most stages of
formed progeny, could not be estimated from these experi-
development, allowing easy selection of transformants.
ments due to the fact that G0 flies were bred in groups.
Secondly, the marker is active in diverse organisms,
However, the overall number of phenotypically detectable
because it depends on the versatile tTA/tetO system rather
transformation events (67 EGFP expressing G1 progeny
than a host-derived promoter. Thirdly, transient expression
from 325 G0 adults) is sufficient to produce several
of the marker in embryos injected with DNA, allowing early
independent transformants from a single experiment, as
assessment of injection efficiency and functionality of the
thousands of embryos can be injected and hundreds of G0
marker. This can be a highly desirable property, especially
adults can be obtained within the productive 10-day period
for transformation of species that have a long generation
time or are difficult to breed. Fourthly, the EGFP expression
Of the nine transformed lines analysed, three lines had a
patterns differ widely between different insertions in some
single copy of the transgene, line 11 carried two copies,
species such as in medfly and in olive fly, a feature that can
lines 10 and 12 three and lines 8, 9 and 13 contained more
be used to distinguish easily between different transgenic
than three copies, contrary to other Minos-based trans-
lines. Additionally, the system can be used as a base for
formation experiments (Loukeris et al., 1995a,b) where most
conditional expression of exogenous genes in various spe-
of the individuals analysed contained a single copy of the
cies. The results presented here suggest that the system
transgene. This difference may be a function of the species
may also be useful as a sensitive enhancer-trap (Bellen
transformed. Alternatively, it may be the result of high levels
et al., 1989; Wilson et al., 1989) for identification of tissue-
of transposase in embryos injected with mRNA, rather than
or stage-specific genes in diverse species.
with a transposase-expressing plasmid (Kapetanaki et al.,
It has been shown recently that a construct expressing
2002). The presence of multiple Minos insertions in lines
tTA from a tetO promoter causes tetracycline-repressible
such as line 13, which show Mendelian inheritance of
lethality in transgenic medfly (Gong et al., 2005); toxicity of
EGFP expression, suggests that all insertions except one
overexpressed tTA was presumably the cause of lethality.
are silent, that is, they do not express EGFP. This is sup-
Such lethals would not have been recovered in our screen
ported by the observation that in subsequent generations
2006 The Royal Entomological Society, Insect Molecular Biology, 15, 95–103
some of the insertions are lost without apparent changes in
EGFP expression patterns. A possible explanation for the
Donor plasmid pMiBO14/GtTA2, containing the ‘insect’ self-
existence of silent insertions is discussed below.
sustaining marker expression cassette was constructed as follows:
Multiple integrations, although useful for enhanced trans-
The EGFP cassette along with the hsp70 minimal promoter and
gene expression, may increase the potential for lethal or
the SV40 polyadenylation sequence was isolated from p3XP3-EGFP (Horn & Wimmer, 2000) as a Bst BI (filled-in)/ClaI fragment
semilethal mutations, decreasing the viability of transgenic
and was cloned into the EcoRV and ClaI sites of pBluescript II SK
lines and furthermore, may complicate applied use of trans-
(+) (Stratagene, La Jolla, CA, USA) to generate pBS / hsTATA-
genic strains. This, however, could be avoided by using
EGFP. The tetracycline operator (tetO) sequence was obtained as
transposase-expressing helper plasmids or lower concen-
an AluI fragment from PBI-L (Clontech, Mountain View, CA, USA).
trations of transposase mRNA in embryo injections.
Two copies of this fragment were cloned into the SmaI site of
Patterns of EGFP fluorescence varied dramatically
pBluescript II SK (+) to give a tetO-14mer (pBS/14tetO). The
among different transgenic lines of olive fly (Fig. 2) and
hsp70 minimal promoter, the tTA gene, the hsp70 polyadenylationsequence and the tetO 14mer (tetO-14) were then cloned into an
medfly (Fig. 1); in these species EGFP expression was
EcoRI/NotI-digested pBS/hsTATA-EGFP in a five-fragment ligation
characteristically tissue-specific and variegated. In con-
giving pBO14/GtTA. The hsp70 minimal promoter was on a
trast, the majority of Drosophila transformants carrying
BamHI/KpnI fragment from plasmid p3XP3-EGFP (Horn & Wim-
single insertions of the self-stimulating cassette exhibited
mer, 2000), hsp70pA was on a XbaI/NotI fragment from plasmid
strong and ubiquitous expression of EGFP (Fig. 1). Expres-
pHSS6hs/LMi2 (Loukeris et al., 1995b) and tetO-14 was on a
sion of the EGFP gene from the self-activating construct
BamHI/NotI fragment from pBS / 14tetO. The tTA gene was a KpnI/
used in these experiments depends on initial activation
NheI fragment from plasmid pBI / GtT. Plasmid pBI / GtT was con-structed as follows: the EGFP gene was cloned on a Pst I/XbaI
(triggering) of the minimal Hsp70 promoter. It is known that
fragment from plasmid pEGFP-N1 pBI/GtT (Clontech) into plasmid
the complete Drosophila Hsp70 promoter exhibits low
pPBI-l (Clontech), replacing the luciferase gene, resulting in plasmid
activity in transgenic non-Drosophila species (Berger et al.,
pPB1-g. The tTA gene derived from pUHD15-1 plasmid (Gossen
1985; our own unpublished results from medfly). It is pos-
& Bujard, 1992) on a EcoRI/BamHI fragment was then cloned
sible that the minimal Drosophila Hsp70 promoter is also
blunt into the PvuII site of pPB1-g, resulting in plasmid pBI / GtT.
less active in medfly and olive fly, failing to ‘trigger’ the self-
The XhoI site of pBO14/GtTA was destroyed by fill in to give
sustaining loop in these species, unless the transposon has
pBO14/GtTA2. An XhoI/XbaI fragment of this vector containing thewhole cassette except for the tTA gene and the hsp70pA was
inserted near an active enhancer. This could explain both
cloned into pMiLRtetR (Klinakis et al., 2000b) to give pMiBO14-G.
tissue-specific, variegated expression from some inser-
The rest of the cassette from pBO14/GtTA2 was subsequently
tions and lack of expression from others.
moved as an XbaI/NotI fragment into pMiBO14-G to reconstitute
This report describes what appears to be a universal
the tTA gene, resulting in pMiBO14/GtTA2. The donor plasmid
genetic marker for detection of transgenic organisms and
pMiBO7/GtTA was constructed in an analogous manner as
its use for transgenesis of the major olive pest B. oleae by
pMiBO14/GtTA2. Minos transposase mRNA was synthesized
means of Minos-mediated germline transformation. With
using the linearized plasmid pBlue(SK)MimRNA as a template(Pavlopoulos et al., 2004), according to the manufacturer’s instruc-
this system, the olive fly is now amenable to transgenesis.
tions (Ambion mMessage mMachine T7 kit Austin, TX, USA).
Availability of a transformation system for B. oleae shouldnow make it possible to use genetic engineering for develop-ment of genetic strategies for control of this important
Expression from the tTA/EGFP cassette in animal and plant cells
Germ line transformation of D. melanogaster and Ceratitis capitatawas performed as described previously (Loukeris et al., 1995a,b). HeLa cells were cultured and transfected with super-coiled plas-
Experimental procedures
mid DNA by Ca++ coprecipitation as described previously (Klinakiset al., 2000b) and examined for EGFP fluorescence 12 h post-
transfection. Transfection of Nicotiana tabacum leaf cells was
The olive fruit flies used in this work originated from the Democri-
carried out with a PDS-1000/Helios Bio-Rad gene-gun (Bio-Rad,
tos Laboratories (Athens, Greece) B. oleae stock. Flies were bred
Hercules, CA, USA). DNA was loaded on 1 µm gold particles.
locally at 25 °C, 45–50% relative humidity, under a 13 h light/11 h
Bombardment was performed under vacuum using 1100 psi rup-
dark cycle and were fed on a diet consisting of 100 g yeast hydro-
ture discs and the leaf-disc-containing Petri dish was placed on the
lysate, 400 g sugar, 30 g egg yolk and 250 mg streptomycin.
top shelf of the apparatus. Leaves were cultivated for two days in
Newly hatched larvae were transferred on to the surface of larval
Murashige and Skoog (SM) medium (Murashige & Skoog, 1962)
medium consisting of 550 ml distilled water, 30 g soy hydrolysate,
and were subsequently examined for EGFP fluorescence. Stand-
0.5 g potassium sorbate, 2 g Nipagin, 20 g sugar, 75 g brewer’s
ard procedures were followed for Caenorhabditis elegans strain
yeast, 30 ml concentrated HCl, 275 g cellulose powder, 20 ml olive
maintenance, crosses and other genetic manipulations (Brenner,
oil and 7.5 ml Tween-80 (Tzanakakis, 1989). Larvae burrowed into
1974). The nematode rearing temperature was kept at 20 °C.
the food and emerged again when ready to pupate. At this stage
Supercoiled plasmid DNA was injected into the gonads of
they were transferred into small plastic boxes for pupation. Pupae
Caenorhabditis elegans N2 animals, together with plasmid pRF4
were kept at 25 °C until adult emergence.
carrying the dominant transformation marker rol-6(su1006) as
2006 The Royal Entomological Society, Insect Molecular Biology, 15, 95–103 Germ line transformation of the olive fruit fly
described (Mello et al., 1992). Transgenic lines were obtained by
a 32P-labelled GFP probe. The probe was a 720 bp BamHI/XbaI
screening the F generation progeny of injected hermaphrodites
fragment containing most of the GFP gene.
for roller animals. Individual transgenic rollers were examined forEGFP fluorescence.
Genomic DNA (5 µg) of transformants was digested with isoMboI
Olive fly embryo collection and microinjection
(Minotech Heraklion, Greece) and restriction fragments were cir-
Adult females were allowed to oviposit on to ceresin wax cones for
cularized by overnight ligation at 16 °C at a final concentration of
30 min. Deposited eggs were collected by rinsing with deionized
200 ng / ml. PCR was performed on the circularized fragments
water and dechorionated for 1 min in 2.5% sodium hypochlorite,
using primers Imio1 (5′-AAGAGAATAAAATTCTCTTTGAGACG-
followed by extensive washes with deionized water. Eggs were
3′) and Imii1 (5′-CAAAAATATGAGTAATTTATTCAAACGG-3′),
aligned on 24 × 50 mm cover slips using double-sided tape, the
followed by a second round of nested PCR with primers Imio2
posterior pole of each egg facing the outer edge of the tape, and
(5′-GATAATATAGTGTGTTAAACATTGCGC3′) and Imii2 (5′-
covered with halocarbon oil (Sigma, St Louis, MO, USA). Transpo-
GCTTAAGAGATAAGAAAAAAGTGACC3′) (Klinakis et al., 2000a).
son plasmid pMiBO14/GtTA2 (400 ng/µl) and helper RNA (100 ng/
PCR conditions were in both cases: 94 °C for 30 s; 58 °C for 30 s;
µl) (Kapetanaki et al., 2002) in injection buffer (1) were coinjected
72 °C for 2 min; 30 cycles. PCR fragments were separated on a
into the posterior pole of eggs, 45 –120 min after oviposition. Fol-
2% agarose gel. Purified fragments were cloned into vector
lowing injection, G0 embryos were kept for 2–3 days in a humidi-
pGEM-T easy (Promega, Madison, WI, USA) and sequenced
fied Petri dish at 25 °C. Hatchlings were placed on larval food and
grown as described above. G0 individuals were back-crossed tothe parental strain. G0 males were group-mated to virgin femaleflies in a standard Bactrocera oleae 30 × 30 × 30 cm cage at a
Acknowledgements
1 : 4 (male : female) ratio; G0 females were group-mated to males
We thank Dr Kriton Kalandidis for his help with plant cell
at a 1 : 1 ratio. G1 eggs were collected daily until no more viable
transformation and Drs Alexandros Kiupakis and Stefan
eggs were produced. Eggs were washed with dH O and incubated
at 25 °C for 48 h in 0.3% propionic acid for hatching. First instar lar-
Oehler for critically reading the manuscript. This work was
vae were transferred to 90 cm Petri dishes containing larval food.
supported by a Greek Secretariat for Research and
G1 individuals were allowed to develop to third larval instar and
Technology PENED grant to M.K., C.R., A.E., and C.S. References
EGFP positive G1 individuals were transferred into small plastic
Allen, M.L., Handler, A.M., Berkebile, D.R. and Skoda, S.R. (2004)
boxes and left to pupate as described above. Single G1s were
PiggyBac transformation of the New World screwworm, Coch-
backcrossed to wild-type flies in small cages (5 × 5 × 10 cm). G1
liomyia hominivorax, produces multiple distinct mutant strains.
males were kept at a ratio of 1 : 6 with females, G1 females at a
Med Vet Entomol18: 1– 9.
ratio 1 : 1 with males. Thin domes of ceresin wax were introduced
Bellen, H.J., O’Kane, C.J., Wilson, C., Grossniklaus, U., Pearson,
into each cage as oviposition surfaces on the third day of adult life.
R.K. and Gehring, W.J. (1989) P-element-mediated enhancer
Eggs were collected from the fifth day of adult life and subsequent
detection: a versatile method to study development in Dro-
collections occurred every other day until the 10th collection. Third
sophila. Genes Dev3: 1288 – 1300.
instar G2 larvae were screened for EGFP expression and the
Berger, E.M., Marino, G. and Torrey, D. (1985) Expression of Dro-
EGFP-positive larvae selected and backcrossed to the parental
sophila hsp70-CAT hybrid gene in Aedes cells induced by heat
strain as above. Three generations of backcrosses of EGFP-
shock. Som Cell Molec Gen11: 371– 377.
positive adults to the parental strain were performed. Subsequently,
Berghammer, A.J., Klingler, M. and Wimmer, E.A. (1999) A univer-
transformed lines were maintained by selection for EGFP positive
sal marker for transgenic insects. Nature402: 370 – 371.
Brenner, S. (1974) The genetics of Caenorhabditis elegans. Genetics77: 71– 94.
Catteruccia, F., Nolan, T., Loukeris, T.G., Blass, C., Savakis, C.,
Kafatos, F.C. and Crisanti, A. (2000) Stable germline trans-
DNA from single adult flies was purified essentially as described
formation of the malaria mosquito Anopheles stephensi. Nature
previously (Laird et al., 1991). Flies were homogenized in 0.25 ml
405: 959 – 962.
50 mM Tris-HCl pH 8.0, 100 mM EDTA, 1% SDS, 100 mM NaCl,
Coates, C.J., Jasinskiene, N., Miyashiro, L. and James, A.A. (1998)
330 µg/ml proteinase K and incubated at 55 °C overnight. Sam-
Mariner transposition and transformation of the yellow fever mos-
ples were treated with a DNAse-free RNAse solution (200 µg/ml)
quito, Aedes aegypti. Proc Natl Acad Sci USA95: 3748 – 3751.
at 37 °C for 1 h. This incubation was followed by two phenol/chlo-
Drabek, D., Zagoraiou, L., deWit, T., Langeveld, A., Roumbaki, C.,
roform extractions and one chloroform extraction. The supernatant
Mamalaki, C., Savakis, C. and Grosveld, F. (2003) Transposi-
was transferred to a fresh tube, DNA was isopropanol precipitated,
tion of the Drosophila hydei Minos transposon in the mouse
washed with 70% ethanol and resuspended in 20 µl sterile ddH O.
germ line. Genomics81: 108 – 111.
DNA concentrations were determined by spectrophotometry at
Economopoulos, A.P. and Zervas, G.A. (1982) The quality prob-
260 nm. Five micrograms of genomic DNA were digested with
lem in olive flies produced for SIT experiments. Invited inter-
EcoRI, and the resulting fragments separated on a 0.8% agarose
national symposium on the SIT and the use of radiation in
gel. After transfer to nylon membranes, blots were hybridized with
genetic insect control. IAEA SIT/PUB592: 357– 368.
2006 The Royal Entomological Society, Insect Molecular Biology, 15, 95–103
Economopoulos, A.P., Artis, N., Zervas, G., Tsitsipis, G., Hanio-
Klinakis, A.G., Zagoraiou, L., Vassilatis, D.K. and Savakis, C.
takis, G., Tsiropoulos, G. and Mamoukas, A. (1977) Control of
(2000b) Genome-wide insertional mutagenesis in human cells
the olive fly, Dacus oleae (Gmelin), by the combined effect of
by the Drosophila moile element Minos. EMBO Reports1:
insecticides and release of gamma sterilized insects. Z AngewEnt83: 201– 215.
Laird, P.W., Zijderveld, A., Linders, K., Rudnicki, M.A., Jaenisch, R.
Ferreira, J.R. and Tainha, A.M. (1983) Organophosphorus insecti-
and Berns, A. (1991) Simplified mammalian DNA isolation pro-
cide residues in olives and olive oil. Pestic Sci14: 167– 172.
cedure. Nucleic Acids Res19: 4293.
Franz, G. and Savakis, C. (1991) Minos, a new transposable ele-
Lorenzen, M.D., Berghammer, A.J., Brown, S.J., Denell, R.E., Klin-
ment from Drosophila hydei, is a member of the Tc-1 like family
ger, M. and Beeman, R.W. (2003) piggyBac-mediated germ-
of transposons. Nucleic Acids Res19: 6646.
line transformation in the beetle Tribolium castaneum. Insect
Gong, P., Epton, M.J., Fu, G., Scaife, S., Hiscox, A., Condon, K.C.,
Mol Biol12: 433 – 440.
Condon, G.C., Morrison, N.I., Kelly, D.W., Dafa’alla, T., Cole-
Loukeris, T.G., Arca, B., Livadaras, I., Dialedtaki, G. and Savakis,
man, P.G. and Alphey, L. (2005) A dominant lethal genetic sys-
C. (1995a) Introduction of the transposable element Minos into
tem for autocidal control of the Mediterranean fruitfly. Nature
the germ line of Drosophila melanogaster. Proc Natl Acad SciBiotechnol23: 453 – 456. USA92: 9485 – 9489.
Gossen, M. and Bujard, H. (1992) Tight control of gene expression
Loukeris, T.G., Livadaras, I., Arca, B., Zabalou, S. and Savakis, C.
in mammalian cells by tetracycline-responsive promoters. Proc
(1995b) Gene transfer into the medfly, Ceratitis capitata, with a
Natl Acad Sci USA89: 5547– 5551. Drosophila hydei transposable element. Science270: 2002–
Grossman, G.L., Rafferty, C.S., Clayton, J.R., Stevens, T.K.,
Mukabayire, O. and Benedict, M.Q. (2001) Germline trans-
Markaki, M., Craig, R.K. and Savakis, C. (2004) Insect population
formation of the malaria vector, Anopheles gambiae, with
control using female specific pro-drug activation. Insect Bio-
the piggyBac transposable element. Insect Mol Biol10: 597– chem Mol Biol34: 131– 137.
Mello, C.C., Kramer, J.M., Stinchcomb, D. and Ambros, V. (1992)
Handler, A.M., Gomez, S.P. and O’Brochta, D.A. (1993) A func-
Efficient gene transfer in C. elegans: extrachromosomal main-
tional analysis of the P-element gene-transfer vector in insects.
tenance integration transforming sequences. EMBO JournalArch Insect Biochem Physiol22: 373 – 384. 10: 3959 – 3970.
Handler, A.M., McCombs, S.D., Fraser, M.J. and Saul, S.H. (1998)
Michel, K., Stamenova, A., Pinkerton, A.C., Franz, G., Robinson,
The lepidopteran transposon vector, piggyBac, mediates ger-
A.S., Gariou-Papalexiou, A., Zacharopoulou, A., O’Brochta,
line transformation in the Mediterranean fruitfly. Proc Natl Acad
D.A. and Atkinson, P.W. (2001) Hermes-mediated germ-line
Sci USA95: 7520 – 7525.
transformation of the Mediterranean fruit fly Ceratitis capitata.
Haniotakis, G., Koryzakis, M., Fitsakis, T. and Antonidaki, A. Insect Mol Biol10: 155 – 162.
(1991) An effective mass-trapping method for the control of
Murashige, T. and Skoog, F. (1962) Revised medium for rapid
Dacus oleae (Diptera: Tephritidae). J Econ Entomol84: 564–
growth and bioessays with tobacco tissue cultures. PhysiolPlant15: 425 – 433.
Hediger, M., Niessen, M., Wimmer, E.A., Dubendorfer, A. and
Pavlopoulos, A., Berghammer, A.J., Averof, M. and Klingler, M.
Bopp, D. (2001) Genetic transformation of the housefly Musca
(2004) Efficient transformation of the beetle Tribolium casta-domestica with the lepidopteran derived transposon piggyBac. neum using the Minos transposable element: quantitative and
Insect Mol Biol10: 113 – 119.
qualitative analysis of genomic integration events. Genetics
Heinrich, J.C. and Scott, M. (2000) A repressible female-specific
167: 737– 746.
lethal genetic system for making transgenic insect strains suit-
Perera, O.P., Harrell, R.A. and Handler, A.M. (2002) Germ-line
able for a sterile-release program. Proc Natl Acad Sci USA97:
transformation of the South American malaria vector, Anophe-les albimanus, with piggyBac/EGFP transposon vector is
Heinrich, J.C., Li, X., Henry, R.A., Haack, N., Stringfellow, L.,
routine and highly efficient. Insect Mol Biol11: 291– 297.
Heath, A.C.G. and Scott, M.J. (2002) Germ-line transformation
Pucci, C. (1990) Valutazione dell’efficacia delle esche proteiche
of the Australian sheep blowfly Lucilia cuprina. Insect Mol Biol
avvelenate per il controllo del Dacus oleae (Gmel): sperimen-
11: 1– 10.
tazione condotta nel triennio 1988 – 90 nell’alto lazio. Frustula
Horn, C. and Wimmer, E.A. (2000) A versatile vector set for animal
Entomologica, N SXIII(XXVI): 173 – 198.
transgenesis. Dev Genes Evol210: 630 – 637.
Rubin, G.M. and Spradling, A.C. (1982) Genetic transformation of
Jasinskiene, N., Coates, C.J., Benedict, M.Q., Cornel, A.J.,
Drosophila with transposable element vectors. Science218:
Rafferty, C.S., James, A.A. and Collins, F.H. (1998) Stable
transformation of the yellow fever mosquito, Aedes aegypti, with
Sasakura, Y., Awazu, S., Chiba, S. and Satoh, N. (2003) Germ-line
the Hermes element from the housefly. Proc Natl Acad Sci
transgenesis of the Tc1/mariner superfamily transposon MinosUSA95: 3743 – 3747.
in Ciona intestinalis. Proc Natl Acad Sci USA100: 7726 – 7730.
Kapetanaki, M.G., Loukeris, T.G., Livadaras, I. and Savakis, C.
Tamura, T., Thibert, C., Royer, C., Kanda, T., Abraham, E., Kamba,
(2002) High frequencies of Minos transposon mobilization are
M., Komoto, N., Thomas, J.L., Mauchamp, B., Chavancy, G.,
obtained in insects by using in vitro synthesized mRNA as a
Shirk, P., Fraser, M., Prudhomme, J.C., Couble, P., Toshiki, T.,
source of transposase. Nucleic Acids Res30: 3333 – 3340.
Chantal, T., Corinne, R., Toshio, K., Eappen, A., Mari, K.,
Klinakis, A.G., Loukeris, T.G., Pavlopoulos, A. and Savakis, C.
Natuo, K., Jean-Luc, T., Bernard, M., Gerard, C., Paul, S., Mal-
(2000a) Mobility assays confirm the broad host-range activity
colm, F., Jean-Claude, P. and Couble, C. (2000) Germline
of the Minos transposable element and validate new transfor-
transformation of the silkworm Bombyx mori L. using a piggyBac
mation tools. Insect Mol Biol9: 269 – 275.
transposon-derived vector. Nature Biotechnol18: 81– 84.
2006 The Royal Entomological Society, Insect Molecular Biology, 15, 95–103 Germ line transformation of the olive fruit fly
Thomas, D., Donnelly, C., Wood, R. and Alphey, L. (2000) Insect
Tzanakakis, M.E. (1995) Entomology, pp. 365–377. University
population control using a dominant, repressible, lethal genetic
Studio Press, Thessaloniki, Greece. (in Greek)
system. Science287: 2474– 2476.
Wilson, C., Pearson, R.K., Bellen, H.J., O’Kane, C.J., Grossnik-
Triglia, T., Peterson, M.G. and Kemp, D.J. (1988) A procedure for
laus, U. and Gehring, W.J. (1989) P-element-mediated
in vitro amplification of DNA segments that lie outside the
enhancer detection: an efficient method for isolating and char-
boundaries of known sequences. Nucleic Acids Res16:
acterizing developmentally regulated genes in Drosophila. Genes Dev3: 1301– 1313.
Tzanakakis, M.E. (1989) Small-scale rearing: Dacus oleae. In Fruit
Zagoraiou, L., Drabek, D., Alexaki, S., Guy, J.A., Klinakis, A.G.,
Flies their Biology, Natural Enemies and Control. Vol. 3B.
Langerveld, A., Skavdis, G., Mamalaki, C. and Grosveld, F. (2001)
(Robinson, A.S. and Hooper, G., eds), pp. 105 – 118, Elsevier,
In vivo transposition of Minos, a Drosophila mobile element, in
mammalian tissues. Proc Natl Acad Sci USA98: 11474–11478.
2006 The Royal Entomological Society, Insect Molecular Biology, 15, 95–103
1. FIGURAS RETÓRICAS BASADAS EN EL SONIDO. Aliteración: Repetición de uno o varios sonidos con una intención expresiva. Ejemplos: "bajo el ala aleve del leve abanico" (Rubén Darío); "En el silencio sólo se escuchaba / un susurro de abejas que sonaba" (Garcilaso de la Vega); "A las aladas almas de las rosas / del almendro de nata te requiero" (Miguel Hern
MARYLAND STATE DEPARTMENT OF EDUCATION Office of Child Care MEDICATION AUTHORIZATION FORM Regulations permit child care providers to give prescription and non-prescription medication to children in care under certain conditions with prior written permission (Section A) from the child’s parent. A separate form is needed for each prescription or non- prescription medication to be admi
 & Zervas, 1982). Availability of a method for olive flytransgenesis is a prerequisite for genetic manipulation ofthis species for development of more effective controlmethods, such as release of insects carrying a dominantlethal allele (RIDL) strategies (Heinrich & Scott, 2000;Thomas et al., 2000) and sensitization of insect populationsto pro-insecticides (SIPP) (Markaki et al., 2004).
& Zervas, 1982). Availability of a method for olive flytransgenesis is a prerequisite for genetic manipulation ofthis species for development of more effective controlmethods, such as release of insects carrying a dominantlethal allele (RIDL) strategies (Heinrich & Scott, 2000;Thomas et al., 2000) and sensitization of insect populationsto pro-insecticides (SIPP) (Markaki et al., 2004).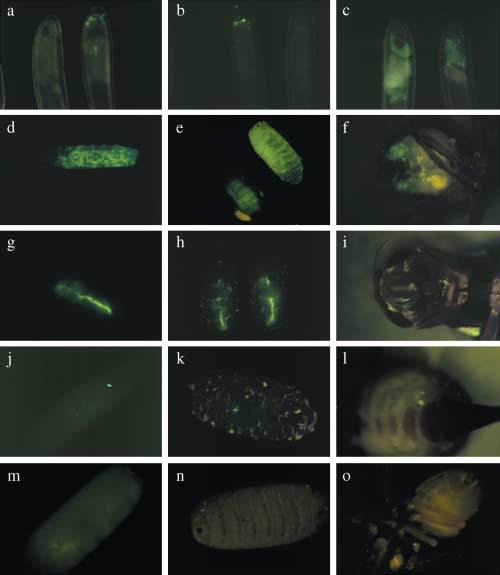 Germ line transformation of the olive fruit fly
The versatility of the bidirectional loop was further
explored in somatic cells of animals and plants. HeLa cells
Olive fly transformation with a Minos element
were transiently and stably transfected with both the
Preblastoderm embryos of the B. oleae Demokritos strain
‘mammalian’ and the ‘insect’ construct; fluorescence was
were coinjected with the Minos-based ‘insect’ construct and
detectable 24 h post transfection (Fig. 1g–h) and peaked
Minos transposase mRNA, a method that has been previ-
48 h post-transfection. The same constructs were also
ously shown to result in high transformation frequencies
introduced into tobacco leaves (Nicotiana tabacum and
(Kapetanaki et al., 2002). Transient EGFP expression was
N. bentamiana) via gold particle ‘bombardment’; examination
observed in about 60% of the injected embryos examined
48 h post transfection showed strong EGFP expression in
(Fig. 2, panels A to C), indicating that the genetic marker is
certain cells (Fig. 1i – j). The ‘insect’ vector was also intro-
duced into the nematode Caenorhabditis elegans, leading
From the 3833 injected G0 embryos, 641 larvae hatched,
to strong and uniform EGFP expression (Fig. 1k).
Germ line transformation of the olive fruit fly
The versatility of the bidirectional loop was further
explored in somatic cells of animals and plants. HeLa cells
Olive fly transformation with a Minos element
were transiently and stably transfected with both the
Preblastoderm embryos of the B. oleae Demokritos strain
‘mammalian’ and the ‘insect’ construct; fluorescence was
were coinjected with the Minos-based ‘insect’ construct and
detectable 24 h post transfection (Fig. 1g–h) and peaked
Minos transposase mRNA, a method that has been previ-
48 h post-transfection. The same constructs were also
ously shown to result in high transformation frequencies
introduced into tobacco leaves (Nicotiana tabacum and
(Kapetanaki et al., 2002). Transient EGFP expression was
N. bentamiana) via gold particle ‘bombardment’; examination
observed in about 60% of the injected embryos examined
48 h post transfection showed strong EGFP expression in
(Fig. 2, panels A to C), indicating that the genetic marker is
certain cells (Fig. 1i – j). The ‘insect’ vector was also intro-
duced into the nematode Caenorhabditis elegans, leading
From the 3833 injected G0 embryos, 641 larvae hatched,
to strong and uniform EGFP expression (Fig. 1k).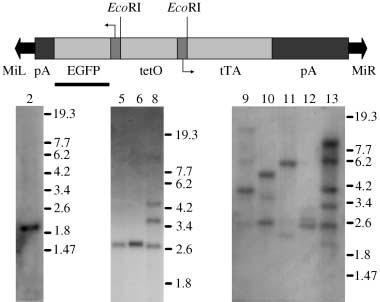 Figure 3. Frequencies of transformants among the olive fly G2 progeny.
Figure 3. Frequencies of transformants among the olive fly G2 progeny.  Germ line transformation of the olive fruit fly
Figure 5. Sequences flanking Minos insertions. The
Germ line transformation of the olive fruit fly
Figure 5. Sequences flanking Minos insertions. The