Tadalafil appartiene alla classe degli inibitori selettivi della fosfodiesterasi di tipo 5, con un profilo farmacocinetico caratterizzato da un’emivita terminale di circa diciotto ore. Dopo somministrazione orale viene assorbito rapidamente e raggiunge concentrazioni plasmatiche massime in due ore. La biotrasformazione avviene principalmente tramite CYP3A4 con formazione di metaboliti inattivi, escreti in prevalenza con le feci. L’elevato legame alle proteine plasmatiche (>90%) assicura una distribuzione stabile. Nei confronti delle altre molecole della stessa classe, cialis compresse italia è noto per la durata prolungata dell’attività farmacologica.
Pdg.cnb.uam.es
Biochemistry I Fall Term, 2001 Lecture 15: Enzyme Inhibition
Assigned reading in Campbell: Chapter 5.7-5.10
Key Terms:
Take the Review Quiz on Lecture 15 concepts:
http://www.bio.cmu.edu/courses/03231/MCQF01/MCQLec15.htm
5.7 Inhibition of Enzymatic Reactions
1. Mechanistic studies to learn about how enzymes interact with their substrates.
2. Role of inhibitors in enzyme regulation.
3. Drugs if they inhibit aberrant or harmful biochemical reactions:
• penicillin, ampicillin, et al.: interfere with the synthesis of bacterial cell walls. • methotrexate - anti-cancer drug that affects DNA metabolism in actively growing cells• viagra - interfers with nitric oxide breakdown (NO is a vasodilator)
4. Understanding the role of biological toxins.
• Arsenate - mimics phosphate esters in enzyme reactions, but are easily hydrolyzed. • Amino acid analogs - useful herbicides (i.e. roundup)• Insecticides - chemicals targeted for the insect nervous system. A. Competitive Inhibition
1. Inhibitor binds to the same site on the enzyme as the substrate.
2. Inhibitor only binds to the free enzyme.
3. Inhibitor is usually structurally very similar to the substrate.
The reaction scheme that corresponds to competitive inhibition is:
The inhibitor reduces the amount of E by the formation of EI complex. The inhibitor does not
affect the ES complex after it has formed. The dissociation constant for the inhibitor is K =
There are two anticipated consequences of this additional competive equilibrium:
• V is unchanged: At high levels of substrate all of the inhibitor is displaced by
• K is increased: Higher substrate concentrations are required to reach the maximal
Steady-state analysis of the effect of the inhibitor shows that K is increased by a factor of
(1 + [I]/K ). The resulting form of the Michaelis-Menten equation is:
Measurements of complete saturation curves (i.e. v versus [S]) at different inhibitor
concentrations can be used to obtain the dissociation constant for inhibitor binding. The
Lineweaver-Burk plots will show an unchanged V
and a slope that increases with inhibitor
Campbell's eqn. (5.18) for the double reciprocal plot shows that the slope of the lines will be:
Thus, K can be determined by plotting the slope values vs. [I]. The resulting secondary plot or
"replot" will have a Y-axis intercept of K /V
The lecture handout, "Data Analysis for Competitive Inhibition", shows an example of the use of
B. Noncompetitive Inhibition:
In this case the inhibitor binds to both E and ES. Both the slope (K /V
) of the Lineweaver-Burk plot increase (see figure 5.11). The K ('s) are determined as
above by replotting the slope and intercept values vs. [I].
• V is decreased: At high levels of substrate the inhibitor is still bound.
• K is increased: Higher [S] is required to reach the lower maximal velocity. (For
"simple noncompetitive inhibition", K is not changed, i.e. Fig. 5.11 in Campbell.)
C. Uncompetitive Inhibition (not mentioned in Campbell)
1. The inhibitor binds directly to the ES complex.
2. The inhibitor does not have to bind at the active site.
3. The inhibitor does not have to resemble the substrate (e.g. allosteric inhibitor). is reduced: the amount of ESI formed depends on [I]. The solution to the steady-state
equation results in the (1 + [I]/K ) factor multiplying the [S] term in the denominator of the
The slopes of the Lineweaver-Burk plot are unchanged (K /V
by a factor of (1 + [I]/K ). The X-intercept shifts to the left by a factor (1 + [I]/K ). Irreversible Inhibition
In contrast to the above types of reversible inhibition, where the effects on the enzyme reaction
depend on the concentration of an inhibitor (and its K ), there are many examples of compounds
that react chemically with residues in the enzyme active site. In these cases, enzyme activity is
destroyed. For example, the "nerve gas" Sarin reacts specifically with an active site Ser residue
on the enzyme, acetylcholinesterase. If acetlycholine cannot be hydrolyzed by this enzyme,
nerve signals cannot be passed across the synapses of the nervous system. On exposure to this
compound, death can result in minutes due to respiratory failure. 5.8 "The Michaelis-Menten equation does not describe the behavior of allosteric enzymes."
The above title statement, quoted from Campbell, is misleading. What is true is that the simplest
form of the Michaelis-Menten equation does not account for the higher than first order substrate
concentration dependence found in many allosteric enzymes. But this is the same as saying that
the Scatchard equation does not describe O binding by hemoglobin. In each case, the basic
concept of saturation (of an enzyme active site or a ligand binding site) is identical. In fact, the
cooperativity observed in allosteric enzymes is analyzed using the Hill equation and Hill plots to
Allosteric enzymes are usually found at metabolic control points. Frequently, these are the first
steps in pathways. Regulation of the entire sequence of enzymatic steps is most efficiently
accomplished by turning on or turning off the pathway at the first step. When the control is
exerted by the end product of the pathway, the inhibition (or activation) is termed "feedback
regulation". An example to be considered later is the feedback inhibition by ATP of the
glycolytic pathway. Allosteric regulation always involves the binding of a metabolite to a site
separate from the enzyme active site. In this regard, the regulation is analogous to the allosteric
effects of BPG on O binding by hemoglobin. 5.9 Models for the behavior of allosteric enzymes
The concerted and sequential models for describing allosteric behavior are discussed in detail by
Campbell. Although the terminology has permeated the biochemical literature and is widely
(over-)used, bear in mind that neither model accurately describes any real enzyme-substrate or
protein-ligand interaction! Stated another way, it has not yet been possible to obtain data of
sufficient accuracy to distinguish between the two models. The most intensely studied system is
hemoglobin, where it now appears that a mixture of the two models may be the best (current)
5.10 Zymogens
Campbell's introduction to the topic of proteolytic processing provides the basic concepts of a
frequently-observed regulatory mechanism in metabolism and in the differentiation and
5.10 - 5.13
We will return to these topics at a later date in the semester (after substitution and elimination
reactions have been treated in Organic Chemistry).
“综合英语”课程第四学期英语阅读训练 材料提供人:张少林 Reading Material 4-7 You probably already know that a viselike pressure in the chest is the most common physical sign of a heart attack. You may even be aware that the body sometimes has the pain of a heart I attack, sending to the neck, jaw or arms. But do you know the second, third or fourth most common sign
SAFETY DATA SHEET according to Regulation (EU) No. 1907/2006 1. IDENTIFICATION OF THE SUBSTANCE/PREPARATION AND OF THE COMPANY/UNDERTAKING : Tetracycline hydrochloride The Old Brickyard NEW ROAD, GILLINGHAM Dorset SP8 4XT UNITED KINGDOM 2. HAZARDS IDENTIFICATION Risk advice to man and the environment Irritating to eyes, respiratory system and skin. 3. COMPOSITION/INFORMATIO
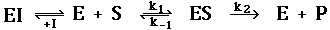
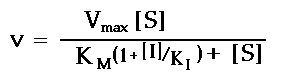
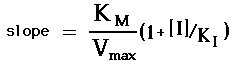 The reaction scheme that corresponds to competitive inhibition is:
The inhibitor reduces the amount of E by the formation of EI complex. The inhibitor does not
affect the ES complex after it has formed. The dissociation constant for the inhibitor is K =
There are two anticipated consequences of this additional competive equilibrium:
• V is unchanged: At high levels of substrate all of the inhibitor is displaced by
The reaction scheme that corresponds to competitive inhibition is:
The inhibitor reduces the amount of E by the formation of EI complex. The inhibitor does not
affect the ES complex after it has formed. The dissociation constant for the inhibitor is K =
There are two anticipated consequences of this additional competive equilibrium:
• V is unchanged: At high levels of substrate all of the inhibitor is displaced by