Tadalafil appartiene alla classe degli inibitori selettivi della fosfodiesterasi di tipo 5, con un profilo farmacocinetico caratterizzato da un’emivita terminale di circa diciotto ore. Dopo somministrazione orale viene assorbito rapidamente e raggiunge concentrazioni plasmatiche massime in due ore. La biotrasformazione avviene principalmente tramite CYP3A4 con formazione di metaboliti inattivi, escreti in prevalenza con le feci. L’elevato legame alle proteine plasmatiche (>90%) assicura una distribuzione stabile. Nei confronti delle altre molecole della stessa classe, cialis compresse italia è noto per la durata prolungata dell’attività farmacologica.
No job name
Biochemistry 1999, 38, 191-198
Tetracycline-Chelated Mg2+ Ion Initiates Helix Unwinding in Tet Repressor
Peter Orth, Wolfram Saenger,* and Winfried Hinrichs
Institut fu¨r Kristallographie, Freie UniVersita¨t Berlin, Takustrasse 6, D-14195 Berlin, GermanyReceiVed July 14, 1998; ReVised Manuscript ReceiVed October 16, 1998
ABSTRACT: The homodimeric tetracycline repressor (TetR) regulates resistance to the antibiotic tetracyclineat the transcriptional level. TetR binds in the absence of Tc to palindromic operator sequences utilizingtwo helix-turn-helix (HTH) motifs. If the tetracycline-Mg2+ complex [MgTc]+ enters two identicalbinding tunnels buried within the TetR homodimer, a conformational change takes place, and the induced[TetR/[MgTc]+]2 complex releases operator DNA. To demonstrate the contribution of Mg2+ to [MgTc]+binding and TetR induction, the Mg2+ concentration in the induced TetR homodimer was progressivelyreduced by addition of EDTA, resulting in two X-ray crystal structures of Mg2+-free and half-occupiedTetR(D). Tc remains bound to the [MgTc]+-binding sites, despite the complete or partial absence ofMg2+. Together with inducer-free TetR(D), the structures were refined to between 2.2 and 2.7 Å resolutionand compared with fully induced TetR(D) in complex with two [MgTc]+. Each inducer binding tunnelhas three constituent parts, one hydrophobic and two hydrophilic ones. One of the hydrophilic contactareas binds Tc by hydrogen bonding; the hydrophobic region correctly positions Tc and partially closesthe entrance to the binding tunnel; the second hydrophilic region coordinates Mg2+, transduces the inductionsignal, and completes the process of closing the tunnel entrance. Tc confers binding specificity to TetRwhile Mg2+ is primarily responsible for induction: After binding to the imidazole N of His100, Mg2+is octahedrally coordinated to the 1,3-ketoenolate group of Tc and to three water molecules. One of thesewaters forms a hydrogen bond to the hydroxyl group Oγ of Thr103. The induced 2.5 Å movement ofThr103 results in the partial unwinding of helix R6, associated with a lateral shift of helices R4 and R9. They simultaneously close the tunnel entrance and cause the DNA-binding domains to adopt a nonbindingconformation, leading to release of operator DNA and expression of the genes responsible for resistance.
The most common resistance mechanism against tetracy-
cline (Tc, Figure 1)1 observed in Gram-negative bacteria isbased on the active efflux of this broad-spectrum antibioticout of the bacterial cell before it can bind to the ribosomeand inhibit protein biosynthesis. The mechanism is regulatedat the level of transcription by the tetracycline repressor,TetR. TetR homodimers bind with their R-helix-turn-R-helix (HTH) motifs to two nearly identical palindromicoperators, tetO1 and tetO2, and block expression of the genestetA and tetR which encode for the membrane-bound Tcefflux protein TetA and the repressor TetR itself (1). Sevenvariants of this resistance determinant are known, referredto as classes A-E, G, and H (2). The polypeptide chains ofTetR contain between 207 and 219 amino acids with at least
FIGURE 1: Chemical structure of the [Mg7ClTc]+ complex, which
occurs under physiological conditions.
† This work was supported by the Deutsche Forschungsgemeinschaft
Due to its high affinity for divalent metal ions (M) under
(SfB 344) and by the Fonds der Chemischen Industrie.
The coordinates for the crystal structures have been deposited in
[MTc]+ (Figure 1) (3, 4). As such, it attacks the TetR/tetO
the Brookhaven Protein Data Bank (codes 1bjy, 1bjz, 1bj0).
* To whom correspondence should be addressed.
complex by binding to TetR with a high affinity of 109 M-1
1 Abbreviations: HTH, helix-turn-helix motif; [MTc]+, complex
(5), associated with a conformational change (induction) of
of a divalent cation, M2+, with Tc; tetA, gene coding for tetracycline
TetR, which lowers the TetR/tetO-binding constant 102-
efflux protein TetA; TetR, Tet repressor; TetR(D), Tet repressor classD; TetR(BD), chimeric construct of TetR(D) and TetR(B); 7ClTc,
103-fold upon binding of the first [MgTc]+ and by an
7-chlorotetracycline; Tc, tetracycline. Crystal forms: [1], native,
additional 104-107-fold after binding of the second [MgTc]+
inducer-free [TetR(D)]2; [2], TetR(D) in complex with two 7ClTc (no
(6). This releases [TetR/[MgTc]+]2 from the operator DNA,
Mg2+; [TetR(D)/7ClTc]2); [3], TetR(D) complexed with two 7ClTc and
and expression of the genes tetR and tetA can now proceed.
one Mg2+ ([TetR(D)/7ClTc‚TetR(D)[Mg7ClTc]+]); [4], TetR(D) com- plexed with two [Mg7ClTc]+ ([TetR(D)/[Mg7ClTc]+]2).
TetA is embedded into the bacterial cytoplasmic membrane
192 Biochemistry, Vol. 38, No. 1, 1999
in the induction of TetR. To shed light on this process, we present two intermediate forms of TetR(D) which were obtained by treating fully induced [TetR(D)/[Mg7ClTc]+]2 [4] with suitable concentrations of EDTA to wholly or partially remove Mg2+ from the complex. In both structures, the two binding tunnels continue to be occupied by 7ClTc. However, while Mg2+ is entirely absent from the inducer binding sites in form [2], [TetR(D)/7ClTc]2, one of the two tunnels retains a Mg2+ in the other, “half-induced” structure [3], [TetR(D)/ 7ClTc‚TetR(D)/[Mg7ClTc]+]. Comparison of these four struc- tures reveals the highly specific roles of both Tc and Mg2+ in the induction mechanism and identifies [MgTc]+ as the actual effector molecule.
To summarize, the following denote the different repressor
states discussed here: form [1], the inducer-free TetR(D) dimer, [TetR(D)]2; form [2], TetR(D) dimer complexed with two 7ClTc (no Mg2+), [TetR(D)/7ClTc]2; form [3], TetR(D)
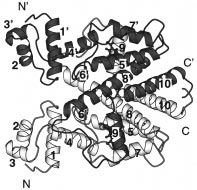
FIGURE 2: Structure of the complex [TetR(D)/[Mg7ClTc]+]2, with
dimer complexed with one 7ClTc and one [Mg7ClTc]+,
one molecule in white and the other in gray. The TetR(D)
TetR(D)/7ClTc‚TetR(D)/[Mg7ClTc]+; and form [4], fully
homodimer is divided into three domains: a large, central inducer-
induced TetR(D) dimer, [TetR(D)/[Mg7ClTc]+]2.
binding core domain and two symmetrically positioned DNA-binding domains. R-Helices 2,3 and 2′,3′ form helix-turn-helix
MATERIALS AND METHODS
motifs. [Mg7ClTc]+ is indicated in ball-and-stick representation. Thisfigure was generated using MOLSCRIPT (23). Crystallization. TetR(D) was overproduced and purified
as described (10). For crystallization, we used the hanging-
and exports invading Tc as [MgTc]+, thereby preventing the
drop vapor-diffusion method at 18 °C. Samples were
ribosomal 30S subunit from being affected because its
prepared by mixing 5 µL of reservoir solution [1 M
affinity to [MgTc]+ is relatively low, 106 M-1 (7). This way
(NH4)2SO4, 200 mM NaCl, and 20 mM Tris-HCl, pH 8.0]
the bacterial cell achieves resistance against Tc.
and 10 µL of the protein solutions described below. El-
The X-ray crystal structure of TetR of class D, TetR(D),
lipsoidal crystals typically appeared within 2-3 weeks and
shows that the homodimer has 2-fold rotational symmetry
grew to dimensions of 0.7 mm × 0.7 mm × 0.3 mm (11).
and consists of three domains: two smaller DNA-binding
To obtain crystals of inducer-free TetR(D) [1], the protein
domains and a large core domain. The polypeptide chain of
solution contained 0.4 mM TetR(D) in 200 mM NaCl, 10
TetR(D) containing 207 residues is folded into 10 R-helices,
mM Tris-HCl, pH 8.0. For crystals of the fully induced
[TetR(D)/[Mg7ClTc]+]2 [4], 0.4 mM TetR(D), 2 mM 7ClTc
Figure 2.) (8). The N-terminal three-helix bundles R -R
(Boehringer Mannheim), 2 mM MgCl2, 200 mM NaCl, and
10 mM Tris-HCl, pH 8.0, were incubated at 30 °C for 30
3 (R2 and R3 , respectively) constitute the HTH motifs.
min and then prepared for hanging-drop vapor diffusion
4 and R4 connect the corresponding DNA-binding
experiments. All Mg2+ was removed from the obtained
domains to the core domain, which is formed by helices R -
crystals by washing them for several days in a reservoir
R10 and their symmetry-related counterparts. The core
solution containing 25 mM EDTA, yielding [TetR(D)/7ClTc]2
domain, as a unit, harbors two identical, symmetry-related
[2]. To obtain TetR(D)/7ClTc‚TetR(D)/[Mg7ClTc]+ [3] (i.e.,
and tunnel-like binding sites for [MgTc]+ formed by side
removing one of the two Mg2+ ions from [4]), 25 mM EDTA
5, R6, R7, R8, R8 , and R9 . Each binding tunnel
was added to the reservoir solution, and mixed with the
has two nonequivalent openings: one of these is flexible
protein solution in a ratio of 1:2 in the crystallization
due to a “sliding door” motion of R ′
experiment, yielding an EDTA concentration in the drop of
represent the entrance for the inducer (9). The other opening
8.3 mM, a 4-fold molar excess over the Mg2+ concentration.
is more rigid and located between R7 and the loop following
X-ray Data Collection. All X-ray data were collected using
R4. After insertion into this tunnel, [MgTc]+ is entirely
the oscillation method (1° rotational increment) and a 180
mm diameter MAR Research imaging plate. Each data set
Relative to the TetR/tetO complex, binding of [MgTc]+
was collected at 4 °C from a single crystal mounted in a
to the TetR tunnel induces a cascade of conformational
glass capillary. X-ray sources included a rotating-anode X-ray
changes within the TetR dimer which culminate in an
generator (Nonius GmbH, type FR571, CuKR radiation,
increased separation of about 5 Å between the HTH motifs,
graphite monochromator, 0.2 × 2 mm2 focal spot size,
located at a distance of 33 Å from the [MgTc]+-binding site.
operated at 45 kV and 65 mA), as well as synchrotron
As a result, the DNA recognition helices R
radiation at the EMBL-outstation (beam line BW7A, DESY,
separated by 39 Å and can no longer bind to adjacent major
Hamburg) and at the SRS Daresbury (station 9.5). The data
grooves of B-DNA (with repeat of 34 Å), so that the operator
were reduced using DENZO (12), scaled with SCALEPACK
(12), and further processed with programs of the CCP4 suite
The inducer-free TetR(D) homodimer [TetR(D)]2 [1] and
(13). Relevant X-ray data statistics for the three data sets
the fully induced [TetR(D)/[MgTc]+]2 [4] represent two
conformational extremes in binding of [MgTc]+ but provide
Structure Solution and Refinement. The previously re-
only a general insight on the specific roles of Tc and Mg2+
ported structure of TetR(D) in complex with 7ClTc and Mg2+
Biochemistry, Vol. 38, No. 1, 1999 193
Table 1: Data Collection and Refinement Statistics
I/σ(I) (last 0.1 Å shell)
FIGURE 3: Difference electron-density maps (coefficients F -
Fc, contoured at 2.5σ) showing 7ClTc and [Mg7ClTc]+ bound to TetR(D).
The maps were calculated using phases derived from models of forms [2] (a), [3a] (b), and [3b] (c), with inducer omitted, using the program X-plor (15). This figure was generated using BOBSCRIPT (24) and RASTER3D (25).
[[4], PDB entry code 2TCT (11)] served as a model in
per asymmetric unit; the local dyad of the homodimeric
molecular replacement for crystal [3], using the program
TetR(D) coincides with a crystallographic 2-fold axis parallel
AMoRe (14). The crystals of the Mg2+-free repressor/Tc
to the 4-fold screw axis. The average temperature factor for
complex [2] were sufficiently isomorphous to [4] so that
main-chain and side-chain atoms is 57.6 Å2. The molecular
atomic coordinates could be used directly for initial phasing.
structure is very similar to that reported for a chimeric
The software package X-plor V3.8 (15) was employed for
inducer-free [TetR(BD)]2 (9) where the DNA-binding domain
refinement of forms [1], [2], and [3]. The 7ClTc geometry
in TetR(D) is replaced by the corresponding polypeptide
was restrained according to parameters similar to those used
sequence of TetR(B) differing only by 10 amino acid
in protein refinement (16). All refinement procedures were
residues. The rms deviation between main-chain atoms of
corroborated by analysis of the free R-factor, and model bias
these two structures is 0.66 Å. The distances between the
was reduced by following a simulated annealing protocol.
The R-factor converged (Table 1) with alternate cycles of
(40.6 Å in [TetR(D)]2 and 40.9 Å in [TetR(BD)]2). The size
manual model building using the program FRODO (17) as
of the entrance to the binding tunnel is 81 Å2, ensuring the
well as atom positional and restrained isotropic B-factor
(B) [2] Mg2+-Free, Tc-Bound TetR(D); [TetR(D)/7ClTc]2.
This Tc-containing form of TetR(D) is obtained from the
fully induced TetR(D) [4] by washing the crystals with a The Four Crystal Structures. Here we describe the
large excess of EDTA which leads to the complete depletion
characteristic features of the individual structures. Together
of Mg2+. The space group (I4122) and the packing arrange-
with the previously determined structure of the induced
ment of TetR(D) dimers remain unchanged as compared to
[TetR(D)/[Mg7ClTc]+]2 (11), they represent four “possible”
[1], but the average B-factor increases to 68.0 Å2 compared
states of complex formation between TetR, Tc, and/or
to 57.6 Å2 for crystals of [1]. The electron density of the
bound 7ClTc corresponds to 50% occupancy, indicating a
(A) [1] NatiVe or Free [TetR(D)]2. This is the inducer-
loose association between TetR(D) and 7ClTc (Figure 3). The
free form of TetR(D) with unoccupied [MTc]+-binding
entrance of the binding tunnel has an area of 72 Å2, which
tunnels. The space group is I4122 with one TetR(D) monomer
is only a little smaller than the 81 Å2 of [1], because helices
194 Biochemistry, Vol. 38, No. 1, 1999
FIGURE 5: Comparison of forms [2] (dark) and [4] (light). Binding of Mg2+ to the inducer in the binding pocket results in C-terminal
FIGURE 4: Conformational change of the entrance to the TetR(D)-
binding tunnel on binding of inducer [Mg7ClTc]+. Structures: long-
6 and the rotation of bound 7ClTc. The residues
His64, Asn82, and Gln116 (group I) which serve to anchor 7ClTc
dashed line, [1] [noninduced TetR(D)]; continuous line, [3a] (half-
through hydrogen bonds to ring A remain largely unchanged. The
induced monomer of [3]); and short-dashed line, [4] (fully induced
side chain of Pro105 points to the upper side of the Tc-rings C and
D and is omitted for clarity; for the same reason, Arg104 is shown
are shifted toward each other, resulting in a narrowing of the
as a thin line. This figure and Figure 6 were generated using SETOR
entrance to the binding tunnel. Removal of Mg2+ from the inducer
binding pocket (structures [2] and [3a]) results in the partial opening of the entrance. The opening of the binding pocket in the free TetR is marked as a point-dashed window. The view is similar as in
Table 2: Structural Differences at the [MgTc]+ Entrance
Figure 2 to the lower binding tunnel at R ′
are each shifted slightly (less than 1 Å) to
accommodate 7ClTc. The helices R4 and R6, which are
important for induction (9), and the DNA-binding helices
3 and R3 are similar in length and orientation with respect
to [1]. Thus, the conformation of form [2] is comparable to (C) [3] Partially Induced TetR(D)/7ClTc‚TetR(D)/[Mg7ClTc]+.
is fixed by hydrogen bonds to the conserved amino acid
If the molar excess of EDTA is limited to 4-fold over the
residues His64, Asn82, Phe86, and Gln116 (Figure 4) (8).
Mg2+ concentration, the partially induced complex [3]
These side chains essentially remain unaffected in their
crystallizes within 2 weeks. The overall crystal packing is
positions. As a result, ring D of 7ClTc is rotated away from
similar to [1] and [2] as are the unit cell constants. However,
the entrance (helix R9) of the binding tunnel by about 1 Å
the complex is now asymmetric because of conformational
in [3a] as compared to [2]. The partial closing of the entrance
differences in the two inducer-binding sites. One binding site
through hydrophobic contacts of ring D with side chains of
contains 7ClTc (denoted [3a]), whereas the other is occupied
by [Mg7ClTc]+ (denoted [3b]) (Figure 3). The dyad is lost,
of 7ClTc in the binding tunnel despite the absence of Mg2+.
and the space group is reduced to the lower-symmetrical
[3a] differs from [1] and [2] in the conformation of R-helix
3212; the crystal asymmetric unit contains the complete
7, because the N-terminal amino acid residues, Glu107 to
TetR(D) dimer complex rather than a TetR(D) monomer as
Tyr110, form a 310-helical turn (see Table 2). The difference
in the case of [1], [2], and [4].
between [2] and [3a] is due to the different orientation of (D) [3a] TetR(D)/7ClTc. The conformation of the uninduced
7ClTc in the binding tunnel. In [3a], substituents of Tc-ring
monomer [3a] (Figure 4, thick lines) is similar to both
C form hydrophobic contacts to Val113 moving the center
inducer-free [1] and Mg2+-deficient TetR(D) [2], with rms
of R7 away from the binding tunnel, resulting in deformation
deviations of main-chain atoms of 0.76 and 0.55 Å as
of the N-terminus of R7, which forms a 310-helix. This
compared to [1] and [2], respectively. These superpositions
deformation is also observed in the fully induced TetR(D)
exclude the ‘sliding door’ R9 at the entrance of the binding
cavity of the induced, second monomer [3b] (see below). (E) [3b] TetR(D)/[Mg7ClTc]+. Compared to the uninduced
The overall structural differences between the Mg2+-free
monomer [3a], several significant structural changes are
monomer [3a] and both [1] and [2] (as indicated by the above
rms values) are due to the flexibility of the protein near the
entrance of the inducer binding tunnel in the uninduced forms
Å toward both the DNA-binding domain and the loop
[1] and [2]. This is also illustrated by the observation that
between helices R6 and R7, closing the entrance of the
7ClTc is located closer to the entrance of the binding tunnel
[MTc]+-binding tunnel (see Table 2). Additional hydrophobic
in [2] as compared to [3a]. 7ClTc rotates within the plane
contacts are formed between side chains of Pro105 and
defined by rings B, C, and D around ring A (Figure 5), which
Biochemistry, Vol. 38, No. 1, 1999 195
domains caused by an extended cooperative hydrogen-
Table 3: Temperature-Factor Distribution of 7ClTc-Rings in Forms [2], [3], and [4]a
bonding network formed by eight H2O molecules between
R4 and R7, whereas the inducer-free TetR(D) has sufficiently
mobile three-helix bundles for tetO recognition due to the
Superposition of all four structures shows that in the core
The average temperature-factor B (Å2) of each 7ClTc-ring was
′5 form a rigid scaffold for [MgTc]+ anchoring (9). The
calculated using the six constituent carbon atoms.
similarity of this feature is indicated by Leu204 of theC-terminal loop adjacent to R10. The main chain torsion
(2) In [3a], the two-turn helix R6 is formed by the
angles of this residue are at the borderline of the allowed
oligopeptide segment Gly96 to Gly102, as also observed in
region in all previously observed TetR(D) structures, but the
form [1] and the Mg2+-deficient form [2]. In the induced
side chain is well-defined in the electron density maps.
monomer [3b], Mg2+ coordination is observed as in [4],
At the “far end” of the [MgTc]+-binding tunnel, where
including the unwinding of the C-terminal turn of R6 for
Tc-ring A is tightly bound by hydrogen bonds to side chains
-turn type II formation by residues 100-103. As a
His64, Asn82, Phe86, and Gln116, a second opening was
consequence, the peptide bond plane formed by Leu101 and
located between R7 and the loop following R4. This rigid
Gly102 is rotated by 180° around the CR-C bond of Leu101.
opening was termed as open backdoor for water release
The main-chain dihedral angles of Leu101 and Gly102 are
during inducer insertion into the binding tunnel (9) and shows
(-46°, -45°; -79°, 11°) and (-50°, 132°; 80°, -2°) in [3a]
no significant conformational changes in all four structures.
and [3b], respectively. DISCUSSION
(3) This conformational change of R6 permits movement
of helix R4 of [3b] toward R6 and results in a decreased
In the TetR(D) homodimer, the two [MTc]+-binding
separation of the DNA recognition helices R
tunnels are separated by about 25 Å, and each is again
Å) in comparison to [1] and [2].
separated from the respective DNA recognition helices by
(4) The higher mobility of 7ClTc due to absence of Mg2+
roughly 33 Å. There is thus no direct interaction between
near the center of the inducer-binding tunnel in [2] and [3a]
the inducer-binding sites themselves or between the binding
as compared to [3b] and [4] is shown by an increase of the
sites and the more distant DNA-binding domains. Modes of
B-factors from 7ClTc-rings A-D (see Table 3).
communication between these entities and the proposed step-
(F) [4] The Fully Induced [TetR(D)/[Mg7ClTc]+]2. As in
by-step model for the mechanism of conformational changes
[1] and [2], crystal form [4] exhibits space group I4122 with
required for induction will be discussed by comparison of
one monomer per asymmetric unit. [Mg7ClTc]+ is held in
the four crystal structures of TetR(D) spanning the range
position in the binding tunnel by a network of hydrogen
from the inducer-free to the induced complex.
bonds and hydrophobic contacts, as described (11). The size
As Tc occurs under physiological pH only as [MTc]+
of the entrance to the binding tunnel is smallest in this
complex (3), the forms [2] and [3] are biologically not
structure and is closed (area ) 44 A2), preventing the escape
relevant but, nevertheless, of crucial importance in analyzing
of [Mg7ClTc]+. The rms deviation for main-chain atoms
the structural properties of TetR(D) in different states of 7ClTc
between [3b] and the monomer in [4] is 0.32 Å (excluding
and/or [Mg7ClTc]+ complexation and consequently in ratio-
nalizing the steps sequentially leading to complete induction.
confirming the induced state of monomer [3b]. Binding of Tc to TetR(D). The entrance to the [MTc]+-
9 observed for [3a] relative to
binding tunnel is located between helix R9 and the interhe-
[1] and [2] above is yet more pronounced in [4]sthe
6∩R7 and R8 ∩R9 (9). In [1], the entrance is
difference between the former two amounting to a mere 60%
sufficiently large for the inducer [Mg7ClTc]+ to enter with
of the total shift of 2.04 Å observed for [4] (see Figure 4).
Tc-ring A head-on to ensure recognition with TetR(D).
The distance between the HTH motifs is 39.6 Å, similarly
Within the binding tunnel, the inducer forms site-specific
showing a decrease as compared to the uninduced [1] and
contacts to charged, hydrophilic, and hydrophobic amino acid
[2] and half-induced [3] structures.
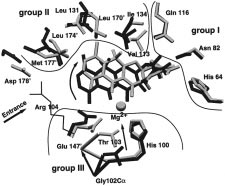
side chains of TetR(D). These may be divided into three
InVariant Features of TetR(D). Besides the [MgTc]+-
categories, all of which have specific functions for inducer
mediated conformational changes, some invariant features
binding and TetR(D) induction (see Figure 5):
in all four structures should be noticed.
(1) Group I is located at the far end of the binding tunnel
In contrast to the significant structural flexibility observed
constituted by the conserved amino acids His64, Asn82,
for the [Mg7ClTc]+-binding tunnels during induction, the
Phe86, and Gln116, whose side chains form hydrogen bonds
structure of the DNA-binding domains does not change.
Notwithstanding this internal rigidity of the three-helix
(2) Group II is the hydrophobic region, coated by type-
3), the distances separating them and their
conserved Val113, Leu131, Ile134, Leu170′, Leu174′, and
relative orientations are critical for tetO binding or release.
Met177′, exclusively involved in nonpolar van der Waals
The differences in the separations of the recognition helices
contacts to the hydrophobic part of [Mg7ClTc]+ with sub-
3, R3 ) in the discussed structures are due to a pendulum-
stituents at positions 6-9 and the corresponding framework
like motion of R4 around the C-terminus. Strictly speaking,
between free and induced TetR, R4 is rotated by 3° according
(3) Group III includes side chains of conserved His100,
His64. The induced state [4] is characterized by a rigid
Thr103, and Glu147′, all involved in direct or water-mediated
4, R4 , and of the associated DNA-binding
196 Biochemistry, Vol. 38, No. 1, 1999The Specific Function of Tc during Induction. A compari-
son of the inducer-free TetR(D) [1] and the Mg2+-free TetR(D)/7ClTc complexes [2] and [3a] allows the specific role of Tc to be investigated, without interference from the complexing metal ion. Clearly, the hydrogen bonds of group I amino acids would be the first to form, after [MTc]+ has entered the binding tunnel. The relative positions of these residues remain essentially unchanged, whether Tc is present or not (Figure 5). Together with the hydrophobic contacts (group II), which guide the Tc to its final position, they serve to recognize and tightly bind the inducer to the binding tunnel.
While the hydrogen bonds and hydrophobic contacts are
formed, the protein segments surrounding the hydrophobicpart of Tc adapt to accommodate and lock the Tc intoposition. The movement of the group II residues induces alateral shift of helix R ′
and R7 form part of the entrance of the binding tunnel, the
hydrophobic contacts serve to partially close the entrance,
Octahedral Mg2+-coordination sphere in the inducer
binding pocket. The ligands are supplied by the two 7ClTc atoms
preventing the Tc from leaving the binding tunnel.
O11 and O12 of the 1,3-ketoenolate, N His100, and three water
Except for the strictly conserved Leu170′, the hydrophobic
molecules W1, W2, and W3. The coordination bonds to Mg2+ are
group II amino acids are only type-conserved in the different
shown as dotted lines; hydrogen bonds to the three water moleculesare dashed lines.
classes of TetR. This agrees with the relative nonspecificnature of hydrophobic contacts allowing Tc analogues with
being provided by three water molecules and the chelating
modified hydrophobic regions to be recognized by TetR and
1,3-ketoenolate of Tc (Figures 1 and 6).
to support induction (18, 19).
A hydrogen-bonding network of the three water ligands
Comparison of the orientations of 7ClTc in forms [2], [3],
of Mg2+ is established with amino acid side chains of Thr103
and [4] shows that the antibiotic occupies two positions (see
and Glu147′. As part of this recognition process, the
Figure 5): an “intermediate” position observed in [2] and
carboxylate group of Glu147′ is rotated by 90°, forming
an “inducing” position seen in forms [3a,b] and [4]. The
hydrogen bonds to two of the Mg2+-coordinated water
two positions differ in rotation of 7ClTc around ring A, which
molecules. The flexibility of the Glu147′ side chain allows
is tightly bound by hydrogen bonds to group I amino acid
this rotation to occur without a significant change in the
side chains found in almost unchanged positions. In the
position of the protein backbone (Figure 5). In hydrogen
“intermediate” position, the long axis of 7ClTc points toward
bonding to the remaining third water molecule, the side chain
the “entrance” and a simple translation parallel to this axis
of Thr103 dramatically shifts by 2.5 Å, forcing the backbone
would cause 7ClTc to leave the binding cavity.
between Gly102 and Arg104 to move by 3.2 Å. Thr103 is
An “intermediate”-like position of the Tc framework was
the first amino acid residue C-terminal to helix R6. Its shift
identified in the crystal structure analysis of TetR(D) in
induces a tension of helix R6 resulting in unwinding the
complex with a 9-(N,N-dimethylglycylamido)-substituted Tc
C-terminal part of R6 to adopt a -turn conformation (residues
analogue (20). This bulky substituent leads to a 10-fold
His100 to Thr103). The amino acid residues of helix R6 itself
decrease of the association constant to TetR (19). The crystal
are in van der Waals contact with those of R4 which is
structure shows that steric hindrance at the entrance and
responsible for positioning of the DNA-binding domain. The
interference with the “sliding door” function of R ′
C-terminal shortening of R6 causes R4 to perform a pendulum-
9-glycyl moiety lead to the “intermediate position” of the
like motion around its C-terminal residue, His64, thereby
Tc framework. As a result, the Mg2+ ion binding to TetR is
communicating the impulse of induction to the DNA-binding
impaired, because the Mg2+-coordination octahedron is not
recognized with the same hydrogen bonding pattern in the
The movement of Thr103 (Figure 5) is accompanied by
binding tunnel of TetR like the fully induced [4] (20).
the neighboring amino acids Arg104 and Pro105, which are
Consequently, in the absence of Mg2+ ions, the binding
conserved in all TetR classes with the exception of Ser104
constant to TetR is about 2 orders of magnitude higher for
in TetR(E). These amino acids are part of the interhelical
9-amino derivatives of Tc (KA ≈ 107 M-1) than for 7HTc
turn between R6 and R7. The functional importance of the
Thr103 side chain is shown by noninducible mutants (21,The Specific Function of Mg2+ during Induction. The
22) where Thr103 is substituted by Ala, Ile, and Lys. In
specific role of Mg2+ in the process of induction is illustrated
addition to the hydrogen bonding to a Mg2+-coordinated
by comparing the uninduced [TetR(D)/7ClTc] structures [2]
water ligand, Thr103Oγ stabilizes the induced -turn by a
and [3a] with the induced [TetR(D)/[MgTc]+] structures [3b]
hydrogen bond to the peptide carbonyl group of His100.
and [4]. Three strictly conserved amino acid residues of
Pro105 as part of the hydrophobic pocket is in contact with
TetR(D) are involved in binding Mg2+: His100, Thr103, and
rings C and D of 7ClTc, which is even tightened due to the
Glu147′. Of these, only N of His100 is involved in the first,
motion of Thr103 leading to a consolidation of the hydro-
octahedral coordination sphere of Mg2+, the other ligands
phobic contacts and a tighter hydrophobic pocket (Figure 4,
Biochemistry, Vol. 38, No. 1, 1999 197
[3b] and [4]). Arg104 in turn forms a salt bridge to the
the DNA-binding domains in the 39.6 Å separation of the
9 (Figure 5), inducing this helix to move
recognition helices R3 and R3′, abolishing the affinity for
yet closer to 7ClTc, thereby reinforcing the shift caused by
operator DNA. The observed interrelation of inducer binding
the hydrophobic contacts between rings C and D to the group
and induction of the TetR/operator complex is an effect of
II amino acid residues. This completes the “sliding-door”
the stepwise changed mobility of the two DNA-binding
and closure of the entrance to the inducer
domains caused by inducer binding. If only one [MgTc]+ is
bound to the TetR(D) dimer, the corresponding HTH motif
Note that the position of 7ClTc is somewhat different in
is forced into the induced orientation and the operator
the Mg2+-free [2] compared to the other states ([3a], [3b],
recognition is significantly weakened. The noninduced
and [4]). Thus, inducer binding proceeds via two steps: First,
monomer is still able to vary its relative position to the other,
insertion of [MTc]+ into the binding tunnel results in
fixed DNA-binding domain and compensates the half-
hydrogen bonding between 7ClTc-ring A and the group I
residues, comparable to that seen in [2]. Second, there is a MoVement of the DNA-Binding Domains. In the four
rotation of 7ClTc around ring A (fixed by its hydrogen-
structures, the separation of the recognition helices R
bonding network) to form the hydrophobic contacts and to
varies between 39.6 Å in the induced state [4] and 40.6 Å
position the chelated Mg2+ appropriate for binding to the
in the uninduced conformation [1]. The apparent ambiguity
with respect to the assumed separation of 34 Å for binding
Only [MgTc]+ is able to trigger the mechanism of
to the major groove of the operator DNA was discussed
conformational changes for induction, but not the Tc
recently (9). In [4], both DNA-binding domains (R -R
molecule without the coordinated Mg2+ ion. This is clearly
positioned by the orientation of R4. After [MgTc]+ binding,
shown by the homodimeric TetR(D) in complex with one
the -turn formation C-terminal to R6 forces R4 into a specific
Tc and one [MgTc]+. If only Tc occupies the binding tunnel,
position, which is fixed by an extending network of water-
the corresponding TetR(D) monomer remains almost un-
mediated hydrogen bonds connecting the inducer binding site
changed and is very similar to the structure of the free
to several peptide carbonyl groups of R4. This ordered water
TetR(D). Recognition of 7ClTc-chelated Mg2+ by imidazole
arrangement was not found in all uninduced structures. The
of His100 allows Thr103Oγ to form a hydrogen bond to a
DNA-binding domains are assumed to be mobile for operator
water ligand of the octahedral Mg-coordination sphere
recognition and separated by their equal charge and probably
initiated by rearrangements of the C-terminal turn of R6 (9).
also crystal packing contacts (9). Communication between the Inducer-Binding Pockets. In
the crystal packing of [3], statistical disorder with respect to ACKNOWLEDGMENT
the induced/noninduced state of the TetR(D) monomers is
We are grateful for synchrotron beam time allocations at
not observed. This is probably due to the crystal contacts of
both the SRS, Daresbury, and the EMBL-outstation at DESY,
the entrance regions of neighboring TetR dimers. The
Hamburg. Our particular thanks are to Heike Roscher for
entrance of an induced monomer is always next to the
preparation of TetR(D) and to Wolf-Dieter Schubert for
entrance of the noninduced half of another TetR dimer. This
systematic packing is probably a more stable arrangementthan the alternative statistical disorder. These crystal packing
REFERENCES
contacts are probably a reason for the observed differentpositioning of R ′
1. Hillen, W., and Berens, C. (1994) Annu. ReV. Microbiol. 48,
9 in the uninduced complexes [2] and [3a]
2. Schnappinger, D., and Hillen, W. (1996) Arch. Microbiol. 165,
The existence of the half-induced TetR(D) with one
monomer in the induced, [MgTc]+-bound conformation and
3. Jogun, H. J., and Stezowski, J. J. (1976) J. Am. Chem. Soc.
the other monomer in the uninduced, Tc-bound conformation
is in agreement with fluorescence-spectroscopic experi-
4. Takahashi, M., Altschmied, L., and Hillen, W. (1986) J. Mol.
ments, where no detectable cooperativity was observed using
5. Takahashi, M., Degenkolb, J., and Hillen, W. (1991) Anal.
free TetR (18, 19). According to our data, conformational
Biochem. 199, 197-202.
changes induced by [MgTc]+ coordination in one binding
6. Lederer, T., Takahashi, M., and Hillen, W. (1995) Anal.
tunnel of the TetR core caused no structural variations in
Biochem. 232, 190-196.
the other one. In contrast, an apparent discrepancy arises from
7. Epe, B., and Woolley, P. (1984) EMBO J. 3, 121-126. 8. Hinrichs, W., Kisker, C., Du¨vel, M., Mu¨ller, A., Tovar, K.,
thermodynamic analysis of inducer binding to the TetR/tetO
Hillen, W., and Saenger, W. (1994) Science 264, 418-420.
complex, where cooperativity was observed. Binding of the
9. Orth, P., Cordes, F., Schnappinger, D., Hillen, W., Saenger,
first inducer already reduces the affinity of TetR to operator
W., and Hinrichs, W. (1998) J. Mol. Biol. 279, 439-447.
by about 3 orders of magnitude. This is sufficient for
10. Ettner, N., Mu¨ller, G., Berens, Ch., Backes, H., Schnappinger,
significant operator release upon binding of the first inducer
D., Schreppel, T., Pfleiderer, K., and Hillen, W. (1996) J. Chromatogr., Sect. A 742, 95-105.
(6) and initiates the resistance mechanism at subinhibitorical
11. Kisker, C., Hinrichs, W., Tovar, K., Hillen, W., and Saenger,
concentrations of Tc (2). The second [MgTc]+ lowers the
W. (1995) J. Mol. Biol. 247, 260-280.
TetR/tetO-binding constant by additional 104-107-fold. The
12. Otwinowski, Z., and Minor, W. (1997) Methods Enzymol. 276,
observed cooperativity can be assigned to the DNA-binding
domains which are operator-bound, whereas in free TetR they
13. CCP4 (1994) Acta Crystallogr. D50, 760-763. 14. Navaza, J., and Saludjian, P. (1997) Methods Enzymol. 276,
After inducer binding to the TetR/operator complex,
15. Bru¨nger, T. A. (1996) X-PLOR Manual Version 3.843, Yale
conformational rearrangements stabilize the orientations of
198 Biochemistry, Vol. 38, No. 1, 1999
16. Engh, R., and Huber, R. (1991) Acta Crystallogr. A47, 392-
22. Mu¨ller, G., Hecht, B., Helbl, V., Hinrichs, W., Saenger, W.,
and Hillen, W. (1995) Nat. Struct. Biol. 2, 693-703.
17. Jones, A. T. (1985) Methods Enzymol., 157-171. 18. Degenkolb, J., Takahashi, M., Ellestad, G. A., and Hillen, W.
23. Kraulis, P. J. (1991) J. Appl. Crystallogr. 24, 946-950.
(1991) Antimicrob. Agents Chemother. 35, 1591-1595.
19. Lederer, T., Kintrup, M., Takahashi, M., Sum, P.-E., Ellestad,
24. Esnouf, R. M. (1997) J. Mol. Graphics 15, 133-138.
G. A., and Hillen, W. (1996) Biochemistry 35, 7439-7446.
25. Merritt, E. A., and Murphy, M. E. P. (1994) Acta Crystallogr.
20. Orth, P., Schnappinger, D., Sum, P.-E., Ellestad, G. A., Hillen,
W., Saenger, W., and Hinrichs, W. (1998) J. Mol. Biol. (inpress).
26. Evans, S. V. (1993) J. Mol. Graphics 11, 134-138.
21. Smith, L. D., and Bertrand, K. P. (1988) J. Mol. Biol. 203,
Premenstrual syndrome (PMS) Overview Premenstrual syndrome (PMS) is a range of physical, psychological and behavioural symptoms that women can suffer up to two weeks before their period begins.1, 2 Doctors are still unsure what causes PMS but it’s been attributed to hormonal change, chemicals called neurotransmitters, hormone-like prostaglandins, diet and lifestyle.2 PMS often increa
FlutiForm® Inhalation Aerosol Confidential and Proprietary Information Synopsis SkyePharma AG Individual Study Table Referring (For National Authority to Part of Dossier: Use Only) Name of Study Drug: Name of Active Ingredient: Fluticasone propionate and formoterol fumarate Title of Study: Long-term Open-label Safety Study with SKP FlutiForm HFA pMDI (100/
 192 Biochemistry, Vol. 38, No. 1, 1999
in the induction of TetR. To shed light on this process, we
192 Biochemistry, Vol. 38, No. 1, 1999
in the induction of TetR. To shed light on this process, we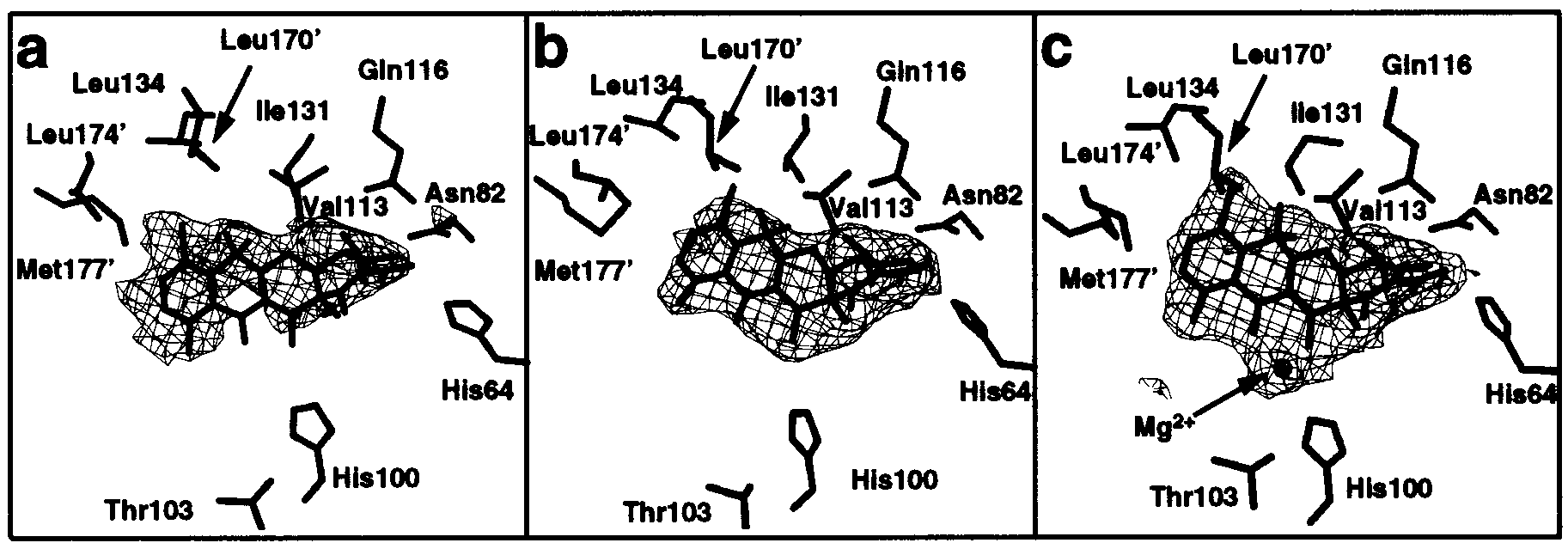 Biochemistry, Vol. 38, No. 1, 1999 193
Table 1: Data Collection and Refinement Statistics
I/σ(I) (last 0.1 Å shell)
FIGURE 3: Difference electron-density maps (coefficients F -
Fc, contoured at 2.5σ) showing 7ClTc and [Mg7ClTc]+ bound to TetR(D).
Biochemistry, Vol. 38, No. 1, 1999 193
Table 1: Data Collection and Refinement Statistics
I/σ(I) (last 0.1 Å shell)
FIGURE 3: Difference electron-density maps (coefficients F -
Fc, contoured at 2.5σ) showing 7ClTc and [Mg7ClTc]+ bound to TetR(D).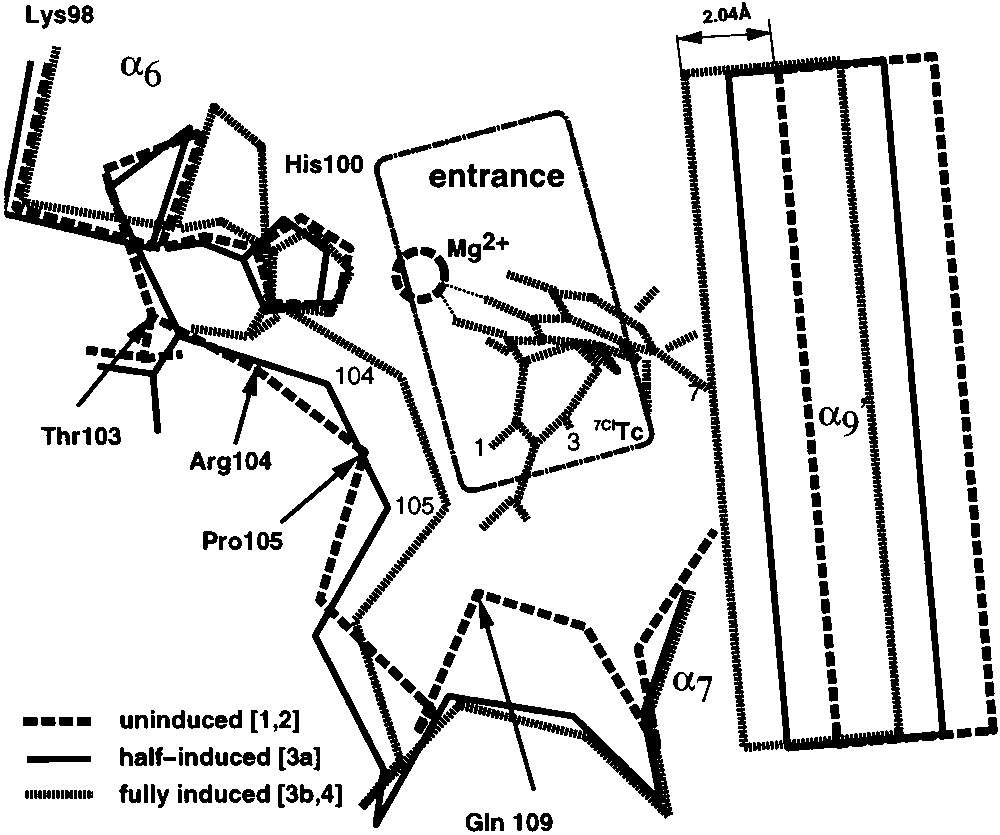
 194 Biochemistry, Vol. 38, No. 1, 1999
FIGURE 5: Comparison of forms [2] (dark) and [4] (light). Binding
194 Biochemistry, Vol. 38, No. 1, 1999
FIGURE 5: Comparison of forms [2] (dark) and [4] (light). Binding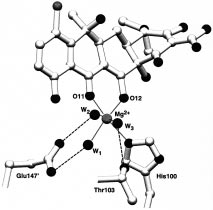 196 Biochemistry, Vol. 38, No. 1, 1999
The Specific Function of Tc during Induction. A compari-
son of the inducer-free TetR(D) [1] and the Mg2+-free
196 Biochemistry, Vol. 38, No. 1, 1999
The Specific Function of Tc during Induction. A compari-
son of the inducer-free TetR(D) [1] and the Mg2+-free