Tadalafil appartiene alla classe degli inibitori selettivi della fosfodiesterasi di tipo 5, con un profilo farmacocinetico caratterizzato da un’emivita terminale di circa diciotto ore. Dopo somministrazione orale viene assorbito rapidamente e raggiunge concentrazioni plasmatiche massime in due ore. La biotrasformazione avviene principalmente tramite CYP3A4 con formazione di metaboliti inattivi, escreti in prevalenza con le feci. L’elevato legame alle proteine plasmatiche (>90%) assicura una distribuzione stabile. Nei confronti delle altre molecole della stessa classe, cialis compresse italia è noto per la durata prolungata dell’attività farmacologica.
Doi:10.1111/j.1600-048x.2008.04632.x
# 2009 The Authors. J. Compilation # 2009 J. Avian Biol.
Received 16 June 2008, accepted 16 November 2008
Test of an adaptive hypothesis for egg speckling along an elevationalgradient in a population of Mexican jays Aphelocoma ultramarina
Elena C. Berg, John E. McCormack and Thomas B. Smith
E. C. Berg (correspondence), J. E. McCormack and T. B. Smith, Center for Tropical Res., Inst. of the Environment, Univ. of California,Los Angeles, La Kretz Hall Suite 300, Box 951496, Los Angeles, USA 90095-1496. E-mail: [email protected]
The adaptive significance of avian egg speckling patterns has been a subject of ongoing debate. We examined speckling ina population of Mexican jays Aphelocoma ultramarina exhibiting extreme eggshell variability. We sampled 167 eggs at 55nests from sites ranging across a steep elevation gradient within the Sierra del Carmen mountain range in Coahuila,Mexico, in order to test the recent hypothesis that egg speckling lends structural support to eggs and should therefore bemore prevalent in females subject to reduced environmental calcium. Although we documented high variation in theamount and distribution of eggshell speckling within the Sierra del Carmen jays, we found no relationship between localsoil calcium levels and the pattern of speckling. Our results indicate that explanations in addition to soil calcium levels arenecessary to explain extreme variation in eggshell speckling in birds.
The wide variety of colours and patterning of the avian
One prediction following from Gosler et al.’s (2005)
eggshell both among and within species has invited con-
hypothesis is that when calcium is limited, speckling should
siderable speculation as to its potential adaptive signifi-
be densest around the crown of the egg, where the shells are
cance. While many hypotheses exist to explain differences in
thinnest (‘‘calcium deficiency hypothesis’’). Gosler et al.
egg appearance, speckling, in particular, is puzzling from an
(2005) found a strong relationship between local soil
adaptive perspective because it occurs in a wide variety of
calcium availability and both eggshell mass and the dis-
species in many ecological contexts Á open-cup nesters,
tribution of speckling on the egg of the great tit Parus
hole-nesters, and species with and without brood parasites
major. Eggshells from low calcium areas were lighter and
(Kilner 2006). In some ground-nesting birds, eggshell
thinner, and speckles were more densely distributed at the
speckling may camouflage the egg and decrease predation
crown of the egg. Thus, while speckling appears to act as
risk (e.g. blackheaded gulls Larus ridibundus, Tinbergen
structural support for eggs of great tits, we do not know if
et al. 1962). However, this hypothesis is less compelling for
the calcium deficiency hypothesis for speckling holds
cup-nesting and hole-nesting species, which are unlikely to
generally for other bird species, and in particular for
receive the same benefits from egg camouflage (Underwood
medium and large birds, where calcium reserves in larger
bones could mitigate environmental calcium deficiencies
Gosler et al. (2005) proposed a novel hypothesis for egg
speckling, suggesting that brown spots, which are produced
Here, we test the hypothesis that egg speckling patterns
by the pigment protoporphyrin and have qualities similar to
are related to environmental calcium available to breeding
solid-state lubricants used in engineering (Solomon 1997),
females of the Mexican jay Aphelocoma ultramarina, a
might provide structural support in areas of the shell that are
medium-sized, cooperatively-breeding passerine bird that
thin or weak. Egg breakage caused by thin or weak eggshells
shows extreme intrapopulation variation in eggshell speckl-
is exacerbated by a deficiency of calcium, which is a limiting
ing. In the Sierra del Carmen of Coahuila, Mexico, where
resource for breeding female birds (Dhondt and Hochachka
we conducted our study, Mexican jays inhabit a steep
2001). The calcium content of soil is highly correlated with
elevation gradient. They are found in highly-divergent
the abundance of calcium-rich invertebrates such as snails,
habitats ranging from arid woodland in the low canyons
and calcium-specific foraging is a common way for many
(1,400 m) to mixed conifer forest at the highest peaks
bird species to supplement calcium levels during the egg-
(2,750 m) (Miller 1955), making small-scale variation in
laying period (Graveland and van der Wal 1996).
of eggshell quality from a limiting-resource perspective, wehave focused exclusively on calcium. For the two nests
Our study was conducted on a population of the couchii
where we collected four samples instead of just one, we also
subspecies of Mexican jays, which ranges from southwestern
conducted an analysis of alkaline earth carbonates (percent
Texas to northern Nuevo Leo´n, Mexico, and is known to
limestone). The first test provides a measure of the amount
have speckled eggs (Ligon and Husar 1974). We collected
of calcium found in organic matter (in parts per million,
data on egg speckling and soil calcium from sites compris-
ppm), while the second test accounts for inorganic sources
ing the full elevational range of Mexican jays in the Sierra
of calcium, such as calcium carbonate, that females might
del Carmen of Coahuila, Mexico, mainly focusing on one
low-elevation oak woodland site (San Isidro Canyon,
All statistical tests were carried out using the statistical
1,458Á1,614 m), one mid-elevation pine-oak-juniper wood-
software package Stata Intercooled (version 10.0, StataCorp
land site (Campo Uno, 1,801Á1,865 m), and two high-
2003). We tested continuous data for normality and
elevation conifer-dominated sites (Campo Dos and Tres,
normalized data when necessary using standard transforma-
2,221Á2,585 m). GPS data including location and elevation
tions. We assessed relationships among variables with
were taken for each nest. Distances between nests ranged
simple regression and multiple regression, using speckling
or speckle clumping as the dependent variable and soil
Using a scalpel, we first confirmed that the speckles on
calcium, elevation, year, clutch size, and interaction terms as
Mexican jay eggs are not superficial markings (as in many
independent variables, removing non-significant terms from
raptor and Charadriiforme eggs; Mikhailov 1997), but that
multiple regression analyses with backward selection. We
they penetrate into the shell. Thus, speckles form part of the
tested for relationships among variables in two ways. First
ground colour and might conceivably lend structural
we assessed the intraclass correlation coefficient for the
support to the shells. We assume that speckles occur at
dependent variables to determine if these variables were
thin parts of the eggshell, as has been shown in other species
highly correlated among eggs within nests. If so, we used
(Gosler et al. 2005, Jagannath et al. 2008). Speckling
nest averages in regressions. Otherwise, we controlled for
percentage and degree of clumping on the crown of the egg
correlations among eggs within clutches with the cluster
were quantified from digital photographs taken using a
(variable) command in Stata, which calculates p-values
Canon PowerShot A40 fitted with a macro lens to prevent
using robust standard errors (Williams 2000). We also
parallax. We used the Java image processing program
analyzed the data with mixed-model ANOVA. Results for
ImageJ (version 1.36b, Abramoff et al. 2004) to quantify
both methods were similar, but we preferred the regression
the area and percentage coverage of speckling on each egg
model because mixed-model ANOVAs can be sensitive
and on each third of each egg: crown, middle, and foot. To
to small sample sizes at the level of the clutch (where 1 B
avoid problems associated with inter-observer variability,
n B5). Means are given9one standard deviation.
only one of us (ECB) conducted all speckling analysis.
We quantified speckling in two ways. First, we calculated
the proportion of the entire (two-dimensional) egg area that
was covered in speckles by dividing the total speckle area bythe total surface area of the egg. Second, we measured the
We collected representative soil samples from 51 of the 55
distribution, or degree of clumping, of speckles across the
nest sites spanning a broad elevation gradient (1,421Á
egg’s surface, per Gosler et al. (2005), using three methods.
2,585 m). Within-nest calcium values were highly corre-
First, we calculated the proportion of the total speckle area
lated (intraclass correlation coefficient 00.97), indicating
that was contained within the crown. Second, to control for
that a single sample is probably sufficient to represent soil
the relative differences in total speckle area across eggs, we
calcium levels in the broader nest area. The two calcium
ran a regression of the crown value against total speckle area
measures Á extractable calcium and calcium carbonates Á
and calculated the residuals. Third, we assigned each egg to
were also highly correlated (Pearson R2 00.72, P 00.044,
one of five categories, ranging from an even speckle
n 08); thus, our analyses focus only on extractable calcium.
distribution (score of 0) to a highly clumped distribution
Across the study site, soil calcium ranged from 783 to
(score of 4). For all analyses of speckling distribution we
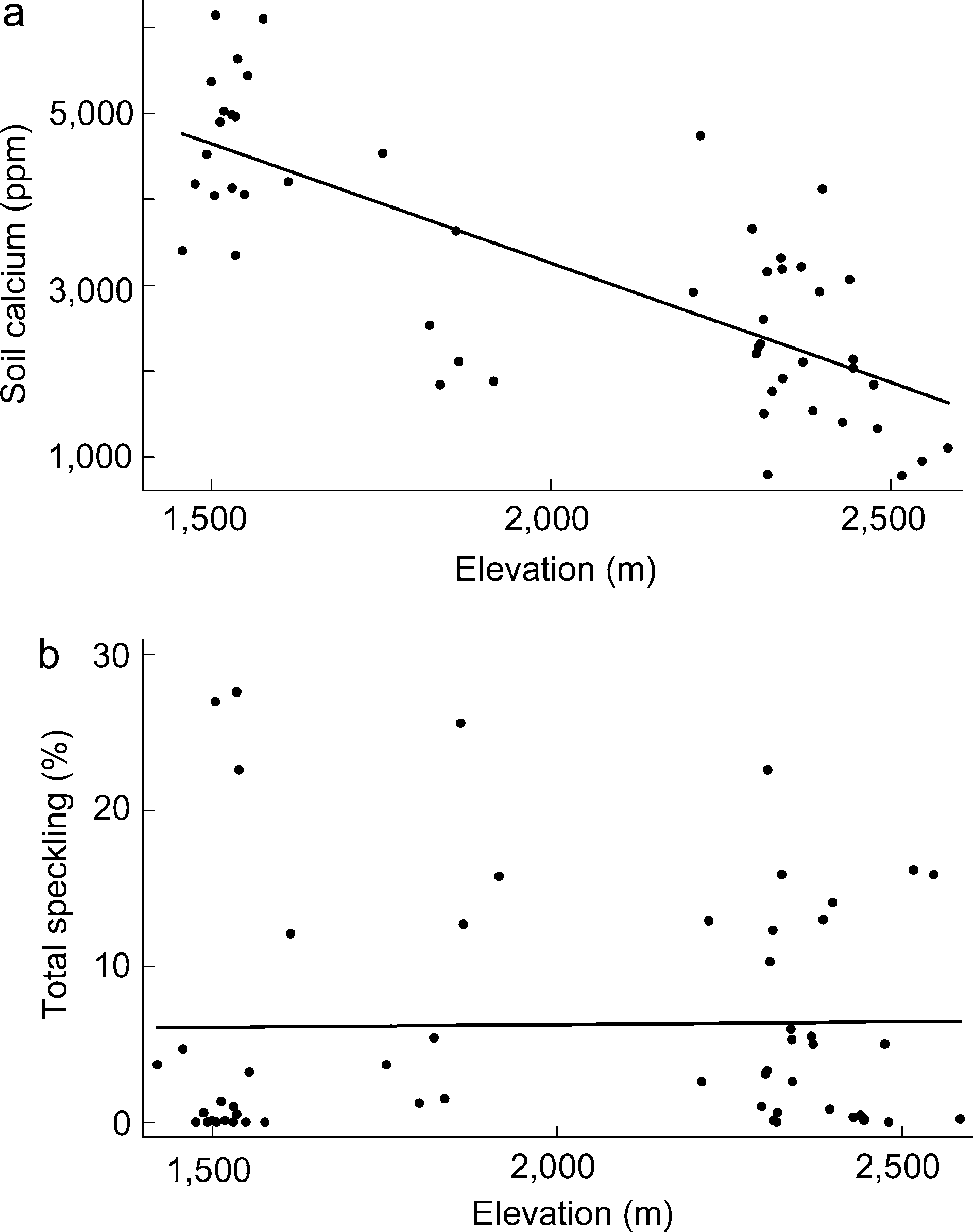
6,138 ppm. There was a highly significant negative relation-
excluded eggs with no speckles (i.e. speckle area of zero).
ship between soil calcium and elevation (regression: F 0
Soil samples of 1 L were collected to a depth of
68.80, df 01,49, P B0.001, R200.58, n 051; Fig. 1a).
approximately six inches from the base of each nest tree.
For example, the lowest-elevation site, San Isidro Canyon
To determine whether soil calcium values varied signifi-
(1,458Á1,614 m), had relatively high soil calcium levels
cantly within the nest area (where the nesting female was
(4,51991,024 ppm, n 020), whereas soil calcium at the
most likely to forage), we collected three additional soil
highest-elevation site, Campo Tres (2,476Á2,585 m), was
samples from randomly-selected locations within a 50-
relatively low (1,1989410 ppm, n 05). Campo Uno
meter radius of the nest tree at one low-elevation and one
(1,801Á1,865 m) and Campo Dos (2,221Á2,400 m) were
high-elevation nest site. Samples were analyzed for extrac-
similar to each other with intermediate calcium levels
table cations, including calcium, at A and L Western
(2,6109712 ppm, n 021 and 2,4889944 ppm, n 05,
Agricultural Laboratories in Modesto, USA. Other minerals
are known to be important to eggshell formation, especially
From 2003Á2007, we measured 167 eggs from 55 nests
in their ratios with calcium (e.g. phosphorus and magne-
along a steep elevation gradient in the Sierra del Carmen.
sium; Fox 1976). However, as these other minerals occur in
From knowledge of flock location, we were reasonably
trace amounts in the eggshell and we approach the question
certain that 42 of these nests were from different flocks and
was no relationship between average speckling per nest(arcsine-square-root transformed to normalize) and eleva-tion (regression: F 00.50, df 054, R2 B0.01, P 00.48,n 055; Fig. 1b), or soil calcium (regression: F 00.49, df 050, R2 00.01, P00.49, n 051).
There was relatively low correlation for speckle clumping
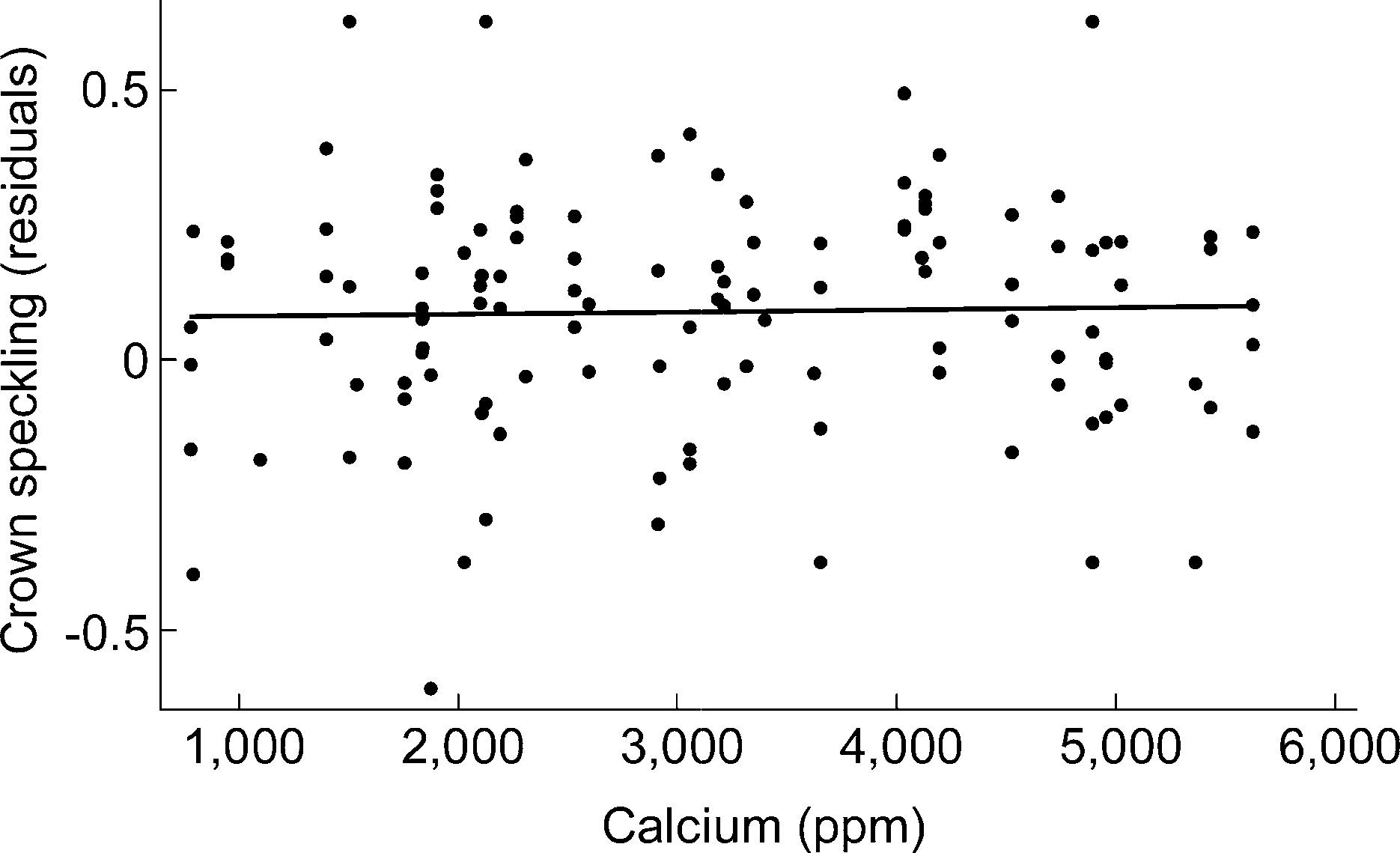
within nests (intraclass correlation coefficient 00.33), so wetreated eggs as individual data points while controlling forcorrelations among eggs within nests (see Methods). Resultsfrom a multiple regression analysis indicated no significantrelationship between speckle clumping and a modelincluding calcium, elevation, year, and calcium)elevation,year)elevation, and year)calcium interactions (regres-sion: F 00.96, df 08,37, R200.05, P 00.46, n 0123). Non-significant terms were removed in stepwise fashionstarting with non-significant interaction terms: calcium )elevation (marginal t 0(0.07; P 00.94), elevation (mar-ginal t 0(1.72; P 00.09) and year )elevation (marginalt 01.93; P00.06), year )calcium (marginal t 01.42;P 00.16), and year (marginal t 0(0.91; P 00.37). Theresulting simple regression between speckle clumping andcalcium was not significant (F 00.08, df01,42, R2 B0.001, P00.78, n 0123; Fig. 2). Even when these datawere analyzed without adjusting degrees of freedom toaccount for non-independence of eggs within nests, therewas no significant relationship between speckle clumpingand calcium (F 00.07, df 01,121, R2 B0.001, P 00.79,
Figure 1. a: Negative relationship between soil calcium and
elevation at nest sites in the Sierra del Carmen (regression: F 068.80, df 01,49, R2 00.58, P B0.001, n 051). b: No relation-ship between average speckling of clutches and elevation (regres-sion: F 00.50, df054, R2 B0.01, P 00.48, n 055).
The population of Mexican jays we studied was particularlywell-suited to a test of the calcium-deficiency hypothesis
therefore highly likely to be from different females. The
because we found high variation in egg speckling (0Á41%
remaining 13 nests were from 10 flocks that were already
surface area speckled) and soil calcium, the latter varying
represented by one nest. Because dominant females often
along an elevation gradient. However, we found that eggs
remain as breeders within flocks over multiple years
from nests at low-calcium sites were not more speckled and
(McCormack and Brown 2008), we were therefore not
speckles were not more clumped toward the crown of the
certain if these 13 nests represented different females. However, because our dependent variables were not highlycorrelated among the nests within a flock (intraflockcorrelation coefficient for speckle clumping 00.36 and forpercent speckling 00.45), we included these nests in ouranalyses. The results did not change when these data wereexcluded.
Speckles ranged from being completely absent to cover-
ing nearly half of the shell’s surface (mean 00.0690.09,range 00 to 0.41). Speckle distribution ranged from beingevenly distributed across the egg to highly clumped towardthe crown, with the proportion of the egg’s speckles locatedon the crown ranging from 0 to 1 (mean 00.4390.29). Our two continuous metrics of speckle clumping werehighly correlated (Pearson r2 00.94, P B0.001, n 0167),and these were correlated with our categorical metric(Pearson r2 00.58, P B0.001, n 073). Thus, we reportonly the results of analyses using the residuals of crown
Figure 2. No relationship between soil calcium and clumping ofspeckles on the crown of the egg (crown speckling) (regression:
speckle area against total speckle area.
F00.08, df01,42, R2 B0.001, P 00.78, n 0123). Values for
Because percentage of the egg’s surface covered with
crown speckling were normalized with arcsine-square-root trans-
speckles (hereafter ‘speckling’) was highly correlated within
formation. Eggs are represented here as independent data points,
nests (intraclass correlation coefficient 00.73), we calcu-
but correlations within nests were taken into account when
lated nest averages for this variable (see Methods). There
calculating the significance of the relationship (see Methods).
egg than at high-calcium sites. Likewise, elevation itself was
tackled by returning to and rigorously testing hypotheses
not correlated with either speckling or speckle clumping.
previously considered unlikely in this species, such as egg
Thus, our results do not support the calcium-deficiency
There are two possible explanations for why we might
have failed to detect a relationship between soil calcium and
Acknowledgements Á We thank CEMEX, the McKinneys, D Roe,
speckling. One is that soil calcium levels in the Sierra del
the Del Carmen Project staff, J Brotman, A Byrd, G Castan˜eda,
Carmen might not be low enough to limit eggshell
E Landay, G Levandoski, E Miller, E Pen˜aloza, and M Starling,
formation in Mexican jays. In our study, extractable
D Parker, UCLA Statistical Consulting, and C Bonneaud.
calcium ranged from 783 to 6,138 ppm. In Oxford’s
A Gosler, T Langen, and A Maklakov commented on drafts. Financial support was provided by a UC Mexus Doctoral
Wytham Woods where Gosler et al.’s (2005) study took
Dissertation Grant and a Sigma Xi Grant-in-Aid-of-Research (to
place, extractable calcium ranged from 410 to 230,000 ppm
JEM) and an American Philosophical Society Franklin Research
(Farmer 1995). While the range of values was more extreme
Grant (to ECB). All field procedures were approved by a UCLA
in Gosler et al.’s (2005) study, the low-calcium values
Animal Use and Care Protocol #2005-126-02.
were similar. Thus, all else being equal, female Mexican jaysfrom low-calcium nest sites in the Sierra del Carmen wereexpected to experience similar levels of calcium deficiency asthe great tits in Gosler et al.’s (2005) study. It is important
to note that the range of calcium values that we recordedis similar to the variation found in other studies that
Abramoff, M. D., Magelhaes, P. J. and Ram, S. J. 2004. Image
demonstrated a strong relationship between soil calcium
processing with Image J. Á Biophotonics Intl. 11: 36Á42.
Dacke, C. G., Arkle, S., Cook, D. J., Wormstone, I. M., Jones, S.,
and eggshell characteristics in birds (e.g. Graveland et al.
Zaidi, M. and Bascal, Z. A. 1993. Medullary bone and avian
1994), suggesting that if soil calcium does indeed influence
calcium regulation. Á J. Exp. Biol. 184: 63Á88.
speckling in this species we would have been able to detect
Dhondt, A. A. and Hochachka, W. M. 2001. Variations in
calcium use by birds during the breeding season. Á Condor
Alternatively, Gosler et al.’s (2005) hypothesis might not
be generally applicable to all bird species. Kilner (2006)
Farmer, A. M. 1995. Soil chemistry change in a lowland English
reasons that a ‘‘hierarchy of selective forces’’ has probably
deciduous woodland 1974Á1991. Á Water Air Soil Poll. 85:
influenced the evolution of eggshell polymorphism, with
crypsis providing the most general explanation for variation
Fox, G. A. 1976. Eggshell quality: Its ecological and physiological
significance in a DDE-contaminated common tern popula-
in eggshell colour and patterning, brood parasitism playing
a secondary role, and finally, the need for structural support
Gosler, A. G., Higham, J. P. and Reynolds, S. J. 2005. Why are
driving variation in just a few species. Medium to large-
birds’ eggs speckled? Á Ecol. Lett. 8: 1105Á1113.
bodied birds might not be as dependent on environmental
Graveland, J., van der Wal, R., van Balen, J. H. and van
calcium during the egg formation stage. For instance, Pahl
Noordwijk, A. J. 1994. Poor reproduction in forest passerines
et al. (1997) showed that great tits cannot store calcium in
from decline of snail abundance on acidified soils. Á Nature
their bones, whereas a study of Japanese quail Coturnix
japonica suggested that larger species do draw on medullary
Graveland, J. and van der Wal, R. 1996. Decline in snail
abundance due to soil acidification causes eggshell defects in
bone as a source of calcium during egg-laying (Dacke et al.
forest passerines. Á Oecologia 105: 351Á360.
1993). Mexican jays are much larger than great tits (mean
Jagannath, A., Shore, R. F., Walker, L. A., Ferns, P. N. and
of 120 g versus 19 g), and female jays may thus be able to
Gosler, A. G. 2008. Eggshell pigmentation indicates pesticide
supplement dietary calcium with bone calcium, which can
contamination. Á J. Appl. Ecol. 45: 133Á140.
be stored throughout the year in anticipation of egg laying.
Kilner, R. M. 2006. The evolution of egg colour and patterning in
A study on a larger-bodied species, the sparrowhawk
Accipiter nisus, showed that speckling was indeed related
Ligon, J. D., and Husar, S. L. 1974. Notes on the behavioral
to eggshell thinning, but that the effect was mediated
ecology of Couch’s Mexican jay. Á Auk 91: 841Á843.
through environmental DDT, which blocks uptake of
Madge, S. and Burn, H. 1994. Crows and jays. Á Princeton Univ.
calcium by the shell gland (Jagannath et al. 2008). While
McCormack, J. E. and Brown, J. L. 2008. Mexican jay
this study supports the idea that speckling can provide
(Aphelocoma ultramarina). Á In: Poole, A. (ed.). The birds of
structural support to calcium-deficient eggs even in a raptor,
North America Online. Cornell Lab of Ornithol., Ithaca.
it did not test whether environmental calcium variation was
Http://bna.birds.cornell.edu/bna/species/118.
an important factor or if large-bodied birds instead draw on
Mikhailov, K. E. 1997. Avian eggshells: an atlas of scanning
medullary sources of calcium during egg-laying.
electron micrographs. Á Brit. Ornithol. Club, UK.
Considering that nearly all New World corvids have
Miller, A. H. 1955. The avifauna of the Sierra del Carmen of
speckled eggs (Madge and Burn 1994), including congeners
Coahuila, Mexico. Á Condor 57: 154Á178.
Pahl, R., Winkler, D. W., Graveland, J. and Batterman, B. W.
of the scrub-jay complex, speckled eggs in the couchii
1997. Songbirds do not create long-term stores of calcium in
subspecies of Mexican jays could represent the retention of
their legs prior to laying: results from high-resolution radio-
an ancestral state that currently has no adaptive value.
graphy. Á Proc. R. Soc. B 264: 239Á244.
Before accepting this explanation, however, the adaptive
Solomon, S. E. 1997. Egg and eggshell quality. Á Iowa State Univ.
value of speckling in this species should probably now be
StataCorp. 2003. Stata statistical software. Á StataCorp LP,
Underwood, T. J. and Sealy, S. G. 2002. Adaptive significance of
egg coloration. Á In: Deeming, D. C. (ed.). Avian incubation,
Tinbergen, N., Broekhuysen, G. J., Feekes, F., Houghton,
behaviour, environment and evolution. Oxford Univ. Press,
J. C. W., Kruuk, H. and Szulc, E. 1962. Egg shell removal
by the black-headed gull, Larus ridibundus L.: a behaviour
Williams, R. L. 2000. A note on robust variance estimation for
component of camouflage. Á Behaviour 19: 74Á117.
cluster-correlated data. Á Biometrics 56: 645Á646.
The following is a list of commonly prescribed drugs covered under the EmblemHealth pharmacy program. To obtain the complete formulary, please visit www.emblemhealth.com . The list is not all inclusive and does not guarantee coverage. We encourage you to ask your doctor to prescribe generic drugs whenever appropriate. PLEASE NOTE: Drugs listed on this document may become non-Preferred if a g
BEHAVIORAL EMERGENCIES & Restraints/Emergency Detention Guidelines SIGNS & SYMPTOMS: OBTAIN HISTORY OF: 3. Recent substance (alcohol and drug) use 5. Recent trauma or illness (esp. fever, infection) PRECAUTIONS: 1. Contact or notify law enforcement EMERGENCY MEDICAL RESPONDER: 1. Ensure scene safety. Leave the scene if patient is or becomes violent. 2. Assess
 was no relationship between average speckling per nest(arcsine-square-root transformed to normalize) and eleva-tion (regression: F 00.50, df 054, R2 B0.01, P 00.48,n 055; Fig. 1b), or soil calcium (regression: F 00.49, df 050, R2 00.01, P00.49, n 051).
was no relationship between average speckling per nest(arcsine-square-root transformed to normalize) and eleva-tion (regression: F 00.50, df 054, R2 B0.01, P 00.48,n 055; Fig. 1b), or soil calcium (regression: F 00.49, df 050, R2 00.01, P00.49, n 051).