Tadalafil appartiene alla classe degli inibitori selettivi della fosfodiesterasi di tipo 5, con un profilo farmacocinetico caratterizzato da un’emivita terminale di circa diciotto ore. Dopo somministrazione orale viene assorbito rapidamente e raggiunge concentrazioni plasmatiche massime in due ore. La biotrasformazione avviene principalmente tramite CYP3A4 con formazione di metaboliti inattivi, escreti in prevalenza con le feci. L’elevato legame alle proteine plasmatiche (>90%) assicura una distribuzione stabile. Nei confronti delle altre molecole della stessa classe, cialis compresse italia è noto per la durata prolungata dell’attività farmacologica.
Untitled
JOURNAL OF INTERFERON & CYTOKINE RESEARCHVolume 00, Number 0, 2011ª Mary Ann Liebert, Inc. DOI: 10.1089/jir.2011.0062
Interleukin-6 in Sepsis and Capillary Leakage Syndrome
Alexander Kru¨ttgen1 and Stefan Rose-John2
Bacterial sepsis is one of the most frequent and dreaded causes of death in intensive care units. According to thecurrent understanding of sepsis, bacterial components activate innate immune responses via pattern-recognitionreceptors that stimulate signaling pathways, thereby leading to activation of NF-kB and the release of cytokines,alarming the organism and coordinating appropriate defense mechanisms. The resulting ‘‘cytokine storm’’ notonly restricts bacterial invasion; it also harms the host by triggering a hemodynamic collapse with a drop inblood pressure, which could lead to death. One of the cytokines released during sepsis is interleukin-6 (IL-6). Originally described as a B-cell–stimulating factor, this cytokine has since been shown to have multiple addi-tional functions. Interestingly, there is emerging evidence of IL-6 trans-signaling in the pathogenesis of sepsis. We review recent findings and discuss whether therapeutic interference with IL-6 trans-signaling may bebeneficial in this important clinical scenario.
ulation cascades (disseminated intravascular coagulation).
Sepsis (etymologically derived from the Greek word
‘‘rwi2‘‘ meaning ‘‘decomposition’’) is a leading cause of
Eventually, systolic hypotension and diffuse vasoconstriction
death in intensive care units. Sepsis is responsible for 9% of
lead to a fatal therapy-refractory ischemia of multiple organs
deaths per year in the United States and Germany, despite
multiple novel therapeutic approaches tested over the past
Usually, sepsis originates from serious infections, such as
several decades (Stearns-Kurosawa and others, 2011). Be-
pneumonia, or by indwelling medical devices, such as in-
cause bacteria predate humans, sepsis probably predates
travenous lines, that undergo colonization by microorgan-
modern humans as well (Baron and others, 2006).
isms (e.g., bacteria), enabling these invasive pathogens to
Operational terms used by clinicians to describe different
access the blood stream. The spectrum of bacteria isolated
stages of sepsis are systemic inflammatory response syn-
from blood cultures of patients with sepsis has changed
drome (SIRS), sepsis, severe sepsis, and septic shock
profoundly over the past decades. Whereas gram-negative
(Stearns-Kurosawa and others 2011). Systemic inflammatory
bacteria (such as Pseudomonas aeruginosa) were the most fre-
response syndrome is caused by increased levels of pro-
quently isolated culprits in the 1970s, gram-positive bacteria
inflammatory cytokines in the blood and is formally defined
(such as Staphylococcus aureus) and fungi (such as Candida
by altered body temperature ( > 38°C or < 36°C), increased
albicans) are becoming increasingly relevant. The rapid
heart rate ( > 90 beats per minute), altered respiratory rate
transmission of plasmid-based antibiotic-resistance genes
( > 20 breaths per minute), and a white blood count > 12,000
among bacteria (Kru¨ttgen and others 2011), the existence of
cells/mm3 or < 4,000 cells/mm3. Interestingly, SIRS may also
virulence genes, and the propensity of certain bacteria to
be caused by noninfectious diseases, such as trauma, pul-
adhere on indwelling catheter surfaces, forming biofilms, are
monary emboli, and myocardial infarction. In contrast, sepsis
contributing to the increases in hospital-acquired infections;
is defined as SIRS plus infection, whereas severe sepsis in-
such infections are of serious concern for clinicians (Seifert
volves sepsis with dysfunction of at least one organ. Finally,
septic shock is severe sepsis with hypotension (systolic blood
One component of the gram-negative membrane is of par-
pressure < 90 mm Hg or mean arterial pressure < 65 mm Hg
amount significance for the pathogenesis of sepsis: the bacterial
after an adequate fluid infusion). Frequent features in septic
endotoxin lipopolysaccharide (LPS). Release of LPS into the
shock are a high cardiac output and a low systemic vascular
circulation triggers a strong systemic pro-inflammatory re-
resistance state associated with diminished myocardial
sponse reminiscent of septic shock. Most interestingly, it is
function. Blood volume is continually lost into the interstitial
actually the host response to LPS (not the intrinsic properties of
space (due to capillary leakage; see below) and intracellular
endotoxin) that induces the potentially lethal consequences.
locations, and blood vessels are clogged by perturbed coag-
This overwhelming response—to which the human species
1Department of Medical Microbiology, RWTH Aachen University Medical Faculty, Aachen, Germany.
2Institute of Biochemistry, Christian-Albrechts-University Kiel, Kiel, Germany.
might be especially prone—is associated with dramaticallyincreased serum concentrations of pro-inflammatory cyto-kines, commonly known as ‘‘cytokine storm.’’ Although LPSsare intrinsic components of the outer membrane of gram-negative bacteria, lipoteichoic acids and peptidoglycans servea similar role in gram-positive sepsis (Schmidt and others2011).
Both LPS and lipoteichoic acids bind to Toll-like receptors
(TLRs), an important family of pathogen-associated molecular-pattern-recognition receptors alerting the innate immune re-sponse system to the presence of dangerous microorganisms. Whereas TLR4 is the primary LPS receptor, TLR2 is regarded asthe receptor for lipoteichoic acids (Beutler and Rietschel 2003). Other TLRs recognize an extensive array of other microbialcomponents from fungi, viruses, and parasites. If pathogensmay escape the extracellular detection by TLRs and manage toinvade into the cytoplasm of their human host cells, theirpresence may still be detected by nucleotide binding oligo-merization domain-like receptors and intracellular TLRs(Stearns-Kurosawa and others 2011).
TLRs trigger intracellular pathways involving the signal-
ing molecules MyD88 or TRIF and leading to the activationof the transcription factors NF-kB and c-Jun N-terminal ki-nase, thereby initiating the transcription of pro-inflammatorycytokine genes (Stearns-Kurosawa and others 2011). One ofthe major NF-kB target genes is the cytokine interleukin-6(IL-6). Intriguingly, the plasma concentration of this keycytokine may rapidly increase dramatically. Plasma levels ofIL-6 normally range between 1 and 5 pg/mL and have been
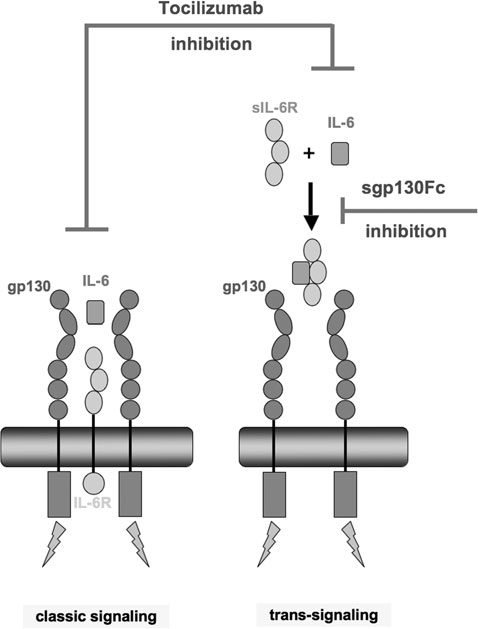
Interleukin-6 (IL-6)/soluble IL-6R (sIL-6) trans-
reported to reach levels > 1 mg/mL during sepsis (Waage
signaling and its inhibition by sgp130Fc. Blood-borne IL-6
and others 1989). Therefore, IL-6 is frequently used as a
and sIL-6R form a complex that binds to and activates gly-
biomarker for sepsis in intensive care units, especially in the
coprotein (gp) 130 at the plasma membrane of target cells
(trans-signaling), whereas classic signaling describes bindingof IL-6 to cells co-expressing gp80 and gp130. Recombinant
soluble sgp130—consisting of the extracellular domain ofgp130 fused with the Fc part of an antibody—acts as a potent
Interleukin-6 is a member of the 4-helical cytokine family,
scavenger by competing with surface membrane gp130 for
which signals via an 80-kDa cytokine receptor (IL-6R, gly-
the binding of the naturally occurring IL-6/sIL-6R complex. Whereas the monoclonal antibody tocilizumab inhibits both
coprotein [gp] 80). Once IL-6 binds to IL-6R, the complex of
classic signaling and trans-signaling, sgp130Fc selectively
IL-6 and IL-6R associates with the signaling receptor subunit
gp130, which thereupon dimerizes and induces intracellularsignaling via the Jak/STAT pathway as well as via the Ras/Map-kinase pathway (Grotzinger and others 1999). Im-
others 1997). The sgp130Fc protein has been used as a mo-
portantly, only few cells in the body—hepatocytes and some
lecular tool to define which gp130-driven murine disease
leukocytes—express IL-6R, whereas gp130 is expressed
models are driven by classic and trans-signaling (Rose-John
ubiquitously (Rose-John and others 2006). Because IL-6 has
and others 2007). It turned out that the pro-inflammatory
no measurable affinity to gp130, it follows that cells, which
activities of IL-6 are mainly driven by IL-6 trans-signaling via
do express gp130 but not IL-6R, are not responsive to IL-6
the sIL-6R, whereas anti-inflammatory or regenerative
(Scheller and others 2011). However, IL-6 signals by 2
functions rely on classic IL-6 signaling via the membrane-
mechanisms: (1) via the ubiquitous transmembrane gp130:
bound receptor (Scheller and others 2011). Of note, it has
‘‘classic’’ signaling using membrane-bound IL-6R (gp80) and
been shown that dying neutrophils, which are the first line of
F1 c (2) via trans-signaling using soluble IL-6R (sIL-6R) (Fig. 1).
defense of the body during infectious diseases, release the
Therefore, cells expressing only gp130 and not IL-6R are
sIL-6R and thereby lead to an attraction of mononuclear
responsive to IL-6 in complex with the sIL-6R (Chalaris and
cells, which are needed for the resolution of the inflamma-
tory process (Chalaris and others 2007).
Even more interestingly, trans-signaling is selectively in-
hibited by soluble gp130. Guided by seminal initial structure-
function analysis of IL-6 (Brakenhoff and others 1989;Kru¨ttgen and others 1990), designer cytokines such as the
Pro-inflammatory cytokines play a pivotal role in the
agonist Hyper-IL6 (consisting of IL-6 fused by a linker to sIL-
pathogenesis of sepsis. Tumor necrosis factor-a (TNF-a) and
6R) and the inhibitor sgp130Fc have been genetically en-
IL-1b are probably the best-examined pro-inflammatory cy-
gineered, adding to our repertoire of tools to elucidate or
tokines. In animal studies, the sole injection of high concen-
manipulate IL-6 signaling in vitro and in vivo (Grotzinger and
trations of TNF-a or IL-1b has lethal consequences (Stearns-
Kurosawa and others 2011). Unexpectedly, the blood levels
of these key cytokines turned out to be of limited use asclinical markers for at-risk patients. Even more disappoint-
Blood vessels are lined with endothelial cells that are in-
ingly, numerous clinical studies failed to demonstrate robust
volved in the regulation of blood pressure. Cell adhesion
clinical benefits from pharmacologic inhibition of TNF-a and
molecules, such as vascular-endothelial (VE) cadherin, form
IL-1b (Stearns-Kurosawa and others 2011). As discussed
cell–cell adherens junctions by connecting neighboring en-
below, this might have been caused by the complexity of
dothelial cell membranes. Clinically, sepsis is associated with
sepsis with—perhaps simultaneously—hyper-inflammatory
tissue edema caused by vascular leakage through endothelial
and anti-inflammatory processes, seriously complicating the
cells (endothelial leakage) that impairs oxygenation of tissues
clear-cut stratification of patients and thereby hampering the
(Lee and Slutsky 2010). Signal transduction pathways trig-
decision about which patients profit at which time point
gered by inflammatory mediators lead to phosphorylation
and endocytosis of VE-cadherin in endothelial cells, giving
Besides IL-1b and TNF-a, a few other interleukins are also
rise to gaps in the endothelial barrier and resulting in serious
of paramount importance. Among these, IL-6 is firmly es-
loss in barrier function. An important recent paper demon-
tablished as a clinically suitable biomarker for sepsis. In
strated the decisiveness of capillary leakage for mortality in
seminal studies, Waage and coworkers (1989) observed high
murine sepsis: London and Li (2011) showed that sealing
levels of IL-6 and its association with fatal sepsis in patients
vascular leaks by infusion of recombinant slit (a soluble
with meningococcal infection. Since then, many other studies
protein best known for its effects in axon guidance of nerve
confirmed and extended these results. For instance, evalua-
cells) completely protected mice from death induced by
tion of postoperative patients with severe sepsis showed that
multiple infectious agents. In line with this, the late Judah
in survivors, IL-6 significantly decreased during the first 2
Folkman demonstrated that therapeutic application of tet-
weeks; in nonsurvivors, IL-6 mostly increased within the
racycline might fulfill a similar role (Fainaru and others
second week (Frink and others 2009; Tschaikowsky and
others 2011). Although IL-6 is an established prognostic
Is there a link between IL-6 and vascular leakage during
marker for mortality in sepsis because of its higher diag-
sepsis? In a visionary paper published in 1997, Ciliberto and
nostic discriminative ability, procalcitonin has overtaken IL-6
coworkers showed that vascular endothelial cells express
in terms of clinical significance (Stearns-Kurosawa and
gp130 but not IL-6R and respond to trans-signaling by IL6/
sIL-6R (Romano and others 1997). In sepsis, considerable
The increased levels of the key cytokine IL-6 correlating
amounts of both IL-6 and sIL-6R are present in the blood
with mortality raised the intriguing hypothesis that IL-6
(Frieling and others 1995), which is separated from organ
signaling plays a mechanistic role in human sepsis. Indeed,
tissues by gp130-expressing vascular endothelial cells. IL-6 is
many studies in mice and cell cultures have corroborated
produced in high amounts upon bacterial infection (Waage
this idea. Remick and coworkers established that measuring
and others 1989), whereas sIL-6R protein is generated by
IL-6 concentrations 6 hours after injury infliction is an ac-
neutrophils, which are likely to be the first cells to encounter
curate predictor of mortality from experimental sepsis in a
invading bacteria (Chalaris and others 2007). Therefore,
mouse model (Remick and others 2002). However, initial
blood-borne IL-6/sIL-6R complexes are bound to interact
work with IL-6 knock-out mice was rather disappointing
with gp130-expressing endothelial cells lining blood vessels.
because it turned out that IL-6 per se might not be of lethal
What are the likely signaling consequences?
importance: These mice had no altered mortality in sepsis
A recent paper from the field of cancer biology provided
(Remick and others 2005). Later on, however, IL-6 signaling
us with a possible answer. Lo and others (2011) showed that
via the gp130 receptor was found to be of paramount im-
IL-6/sIL-6R signaling on human umbilical vascular endo-
portance because mice lacking this receptor in the liver were
thelial cells triggered phosphorylation and redistribution of
protected against an infection with the common sepsis
VE-cadherin, leading to vascular leakage. Because human
pathogen Streptococcus pyogenes (Klein and others 2007). In
umbilical vascular endothelial cells are a standard model for
line with these results, gp130 (F/F) knock-in mice expressing
the study of endothelial cells in sepsis, one effect of IL-6
a gp130 receptor variant with an overactive STAT3 response
trans-signaling in sepsis might be a capillary leakage syn-
were hypersensitive to LPS-induced sepsis (Greenhill and
Fig. 2). This term describes the breakdown of the
others 2011). Moreover, it was shown that trans-signaling in
vascular-endothelial barrier function that normally regulates
vascular endothelial cells seems to play a major role in sepsis
the delicate distribution of nutritional compounds, fluids,
because IL-6 trans-signaling in these cells modulated TLR4-
and leukocytes between blood and tissues. Increased endo-
dependent inflammatory responses (Greenhill and others
thelial permeability by disruption of VE-cadherins function
results in trans-endothelial flow of fluid and interstitial
During shock, the refractory drop in blood pressure is a
edema. This dramatically impairs tissue oxygenation due to
dire clinical problem. Interestingly, a link between IL-6 sig-
increased viscosity of the blood and increased tissue pres-
naling and blood pressure was established; peripheral va-
sure, contributing to therapy-refractory shock (Stearns-
sodilatation in human patients is most strongly associated
with increased IL-6 levels in blood (Hartemink and Groe-
Besides the unexpected role of IL-6 in triggering vascular
neveld 2011). In addition, another interesting study found
leakage in vascular endothelial cell, another recent study
that serum IL-6 correlated with endothelial dysfunction
provided an equally important rationale for interfering with
(Esteve and others 2007). Therefore, an emerging question
IL-6 trans-signaling in sepsis to increase survival outcome
was the mechanism by which IL-6 signaling decreased blood
(Barkhausen and others 2011). Intestinal epithelial cells pro-
pressure and endothelial function, and whether there was a
vide a primary physical barrier against bacteria, and the in-
relation to the development of septic shock.
tegrity of this barrier is damaged when pathologic events
interleukin-6 (IL-6)/soluble IL-6R (sIL-6R) trans-signaling on vascular endo-thelial
Normally, adherens junctions betweenneighboring endothelial cells do notpermit uncontrolled influx of fluid fromthe blood into tissues. During sepsis,IL-6/sIL-6R trans-signaling triggersthe disassembly of these adherensjunctions by inducing the phosphory-lation of VE-cadherin. Influx of bloodinto tissues leads to edema, therebycontributing
lead to a drop in blood pressure with ensuing necrosis. This
Numerous explanations have been brought forward for
is a very serious consequence of shock-induced ischemia: The
resulting necrosis of the gut–blood barrier leads to an irre-
First, interfering with a complex system is complex
sistible invasion of the countless gut-resident commensal
bacteria, to which resistance is futile. Interestingly, Bar-
Second, lack of appropriate stratification of patients might
khausen and others (2011) recently showed in a sepsis model
have played a role in disappointing statistical effects; sepsis
that blocking IL-6 trans-signaling using sgp130Fc blocked
encompasses many different entities that have different
intestinal epithelial cell apoptosis in the gut and decreased
prognoses and different therapeutic needs at different time
mortality. Global blockade of IL-6 by a monoclonal antibody
points (Bone 1995). Support for ‘‘personalized sepsis medi-
had no beneficial effect. Thus, by keeping enteric epithelial
cine’’ is based on the finding that blunting inflammation
cells alive, blocking IL-6 trans-signaling protects patients
improves survival only of animals at a high risk of dying,
with sepsis by keeping the important barrier to enteric bac-
whereas low-dose glucocorticoids are effective in patients
teria sealed, protecting from a ‘‘second front’’ against those
with adrenal malfunction (Remick 2007). The need for a
opportunistic pathogens that threaten to invade the hosts’
tailored therapy in sepsis is already indicated by the obser-
vation that IL-6 levels are significantly higher in gram-negative bacteremia than in gram-positive bacteremia
(Alexandraki and Palacio 2010). Thus, if IL-6 indeed turnsout to be a valid therapeutic target in bacterial sepsis, the
Studies guided by a detailed understanding of the structure–
microbiological origin of sepsis surely needs to be taken into
function relationships of IL-6 with its receptors led to the
account, most likely resulting in differential dosing of IL-6
development of potent IL-6 inhibitors, such as monoclonal
antagonists according to the Gram state of the invading
neutralizing antibodies against IL-6 and its gp80 receptor, as
well as a soluble gp130 Fc fusion protein that inhibits IL-6/sIL-
Third, too late is too late: As a result of therapeutic in-
6R trans-signaling (Brakenhoff and others 1990; Brakenhoff
terference with first-line cytokines during the initial phase of
and others 1994; Grotzinger and others 1997). Previous studies
pathogenesis, therapy may have no benefit when initiated
that applied IL-6 inhibitors in murine disease models of ar-
too late (e.g., during the subsequent late phases of the clinical
thritis and colitis were encouraging and paved the way to
course when patients enter the hospital). Indeed, prospective
clinical studies (Rose-John and others 2007). As a result of such
studies on patients presenting with sepsis showed that in
persistent efforts, tocilizumab, a humanized monoclonal anti-
most patients, peak levels of key cytokines were present on
body that acts as an IL-6 receptor antagonist, has been ap-
the day of admission and decreased on the following days
proved for the treatment of rheumatoid arthritis (Mima and
(Damas and others 1997). Thus, the best time point for in-
terfering with first-line cytokines, such as IL-1b and TNF-a,
Although this seems to be good news for clinicians hoping
might have been missed. Encouragingly, IL-6 levels remain
that IL-6 antagonists will also be approved for the treatment
high and stable during the course of sepsis (Damas and
of sepsis, caution is warranted. So far, the history of the
others 1997). The study by Barkhausen and others (2011)
search for drugs to treat sepsis is so discouraging that this
demonstrated that a therapeutic application of the sgp130Fc
field has even been called the ‘‘pharmaceutical graveyard’’
protein 24 hours after the onset of the sepsis model still
(Riedemann and others 2003). Many of these failed endeav-
ors were aimed at interfering with cytokines known to exert
Fourth, there is a lack of appropriate animal models; mice
detrimental effects in sepsis. Why did so many previous
are mice, and humans are humans. As a species, humans are
expeditions—based on good basic science—eventually fail?
known to be particularly susceptible to lethal effects LPS.
In addition, regarding IL-6, what could still hold up the view
Thus, identification of mechanisms in animals and pharma-
that this time (i.e., with research on modulation of IL-6 trans-
ceutical blockade of these pathways might not translate into
signaling) might be different and that these studies could
clinical efficacy in septic humans (Rittirsch and others 2007).
lead to a panacea for this lethal disease?
Most dramatically, the shortcoming of animal models for
human sepsis became evident in human volunteers in a
not affected by deletion of amino acids 1–28. J Immunol
phase I study of an experimental anti-CD28 antibody (Sun-
tharalingam and others 2006); although this antibody was
Brakenhoff JP, Hart M, De Groot ER, Di Padova F, Aarden LA.
well tolerated in mice and nonhuman primates, all human
1990. Structure-function analysis of human IL-6. Epitope
volunteers given this antibody developed a cytokine storm
mapping of neutralizing monoclonal antibodies with amino-
and shock within a few hours, eventually leading to multi-
and carboxyl-terminal deletion mutants. J Immunol 145:561–
Chalaris A, Garbers C, Rabe B, Rose-John S, Scheller J. 2011. The
soluble Interleukin 6 receptor: generation and role in inflam-mation and cancer. Eur J Cell Biol 90:484–494.
Bacterial sepsis is highly fatal, and its pathophysiology is
Chalaris A, Rabe B, Paliga K, Lange H, Laskay T, Fielding CA,
complex. There is a great need for novel treatments. Suc-
et al. 2007. Apoptosis is a natural stimulus of IL6R shedding
cessful treatment probably requires biomarker-based strati-
and contributes to the proinflammatory trans-signaling func-
fication of patients with sepsis into subgroups, resulting in
tion of neutrophils. Blood 110:1748–1755.
some degree of personalized sepsis medicine. One of the
Damas P, Canivet JL, de Groote D, Vrindts Y, Albert A, Fran-
most relevant biomarkers of sepsis is the multifunctional
chimont P, Lamy M. 1997. Sepsis and serum cytokine con-
cytokine IL-6. Many studies suggest that IL-6 is not only a
centrations. Crit Care Med 25:405–412.
suitable marker but also a key actor in the molecular cho-
Esteve E, Castro A, Lopez-Bermejo A, Vendrell J, Ricart W,
Fernandez-Real JM. 2007. Serum interleukin-6 correlates with
reography of sepsis pathogenesis. Most recently, IL-6 trans-
endothelial dysfunction in healthy men independently of in-
signaling was found to increase endothelial permeability by
sulin sensitivity. Diabetes Care 30:939–945.
phosphorylation of VE-cadherin. We consider this finding,
Fainaru O, Adini I, Benny O, Bazinet L, Pravda E, D’Amato R,
emerging from the field of cancer biology, as very important
Folkman J. 2008. Doxycycline induces membrane expression
for the sepsis field: Because of spatial considerations, vas-
of VE-cadherin on endothelial cells and prevents vascular
cular endothelial cells are bound to be the most likely
hyperpermeability. FASEB J 22:3728–3735.
physiologic target of blood-borne IL-6/sIL-6R complexes.
Frieling JT, van Deuren M, Wijdenes J, van der Meer JW,
Thus, vascular leakage caused by IL-6 trans-signaling (re-
Clement C, van der Linden CJ, Sauerwein RW. 1995. Circu-
sulting in tissue edema and hypoxia) represents a valuable
lating interleukin-6 receptor in patients with sepsis syndrome.
novel target for pharmaceutical intervention and basic sci-
ence. Despite the hope that this novel opportunity will be
Frink M, van Griensven M, Kobbe P, Brin T, Zeckey C, Vaske B,
fruitful, disappointing experiences with different cytokines
et al. 2009. IL-6 predicts organ dysfunction and mortality in
lower our expectations and prompt caution regarding the
patients with multiple injuries. Scand J Trauma Resusc Emerg
value of animal study–derived concepts for human immu-
nology. Nevertheless, the novel IL-6 trans-signaling inhibitor
Greenhill CJ, Rose-John S, Lissilaa R, Ferlin W, Ernst M, Hertzog
is a welcomed addition in our long quest for a much-needed
PJ, et al. 2011. IL-6 trans-signaling modulates TLR4-dependent
inflammatory responses via STAT3. J Immunol 186:1199–1208.
Grotzinger J, Kernebeck T, Kallen KJ, Rose-John S. 1999. IL-6
type cytokine receptor complexes: hexamer, tetramer or both?
S.R.-J. was supported by the Deutsche Forschungsge-
meinschaft (SFB 877, TP A 1) and by the Cluster of Excellence
Grotzinger J, Kurapkat G, Wollmer A, Kalai M, Rose-John S.
‘‘Inflammation at Interfaces.’’ S.R.-J. is inventor on patents
1997. The family of the IL-6-type cytokines: specificity and
describing the function of sgp130Fc and is a shareholder of
promiscuity of the receptor complexes. Proteins 27:96–109.
the CONARIS Research Institute (Kiel, Germany).
Hartemink KJ, Groeneveld AB. 2010. The hemodynamics of
human septic shock relate to circulating innate immunity
factors. Immunol Invest 39:849–862.
Klein C, Medina E, Sander L, Dierssen U, Roskams T, Mueller
Alexandraki I, Palacio C. 2010. Gram-negative versus Gram-
W, et al. 2007. Contribution of interleukin-6/gp 130 signaling
positive bacteremia: what is more alarmin(g)? Crit Care 14, 161.
in hepatocytes to the inflammatory response in mice infected
Barkhausen T, Tschernig T, Rosenstiel P, van Griensven M,
with Streptococcus pyogenes. J Infect Dis 196:755–762.
Vonberg RP, Dorsch M, et al. 2011 Selective blockade of
Kru¨ttgen A, Razavi S, Imohl M, Ritter K. 2011. Real-time PCR
interleukin-6 trans-signaling improves survival in a murine
assay and a synthetic positive control for the rapid and sen-
polymicrobial sepsis model*. Crit Care Med.
sitive detection of the emerging resistance gene New Delhi
Baron RM, Baron MJ, Perrella MA. 2006. Pathobiology of sepsis:
Metallo-beta-lactamase-1 (bla (NDM-1)). Med Microbiol Im-
are we still asking the same questions? Am J Respir Cell Mol
Kru¨ttgen A, Rose-John S, Dufhues G, Bender S, Lutticken C,
Beutler B, Rietschel ET. 2003. Innate immune sensing and its
Freyer P, Heinrich PC. 1990. The three carboxy-terminal
roots: the story of endotoxin. Nat Rev Immunol 3:169–176.
amino acids of human interleukin-6 are essential for its bio-
Bone RC. 1995. Sepsis and controlled clinical trials: the odyssey
logical activity. FEBS Lett 273:95–98.
continues. Crit Care Med 23:1313–1315.
Lee WL, Slutsky AS. 2010. Sepsis and endothelial permeability.
Brakenhoff JP, de Hon FD, Fontaine V, ten Boekel E, Schooltink
H, Rose-John S, et al. 1994. Development of a human
Lo CW, Chen MW, Hsiao M, Wang S, Chen CA, Hsiao SM,
interleukin-6 receptor antagonist. J Biol Chem 269:86–93.
et al. 2011. IL-6 trans-signaling in formation and progres-
Brakenhoff JP, Hart M, Aarden LA. 1989. Analysis of human IL-
sion of malignant ascites in ovarian cancer. Cancer Res 71:
6 mutants expressed in Escherichia coli. Biologic activities are
London NR, Li DY. 2011. Robo4-dependent Slit signaling sta-
Schmidt RR, Pedersen CM, Qiao Y, Zahringer U. 2011. Chemical
bilizes the vasculature during pathologic angiogenesis and
synthesis of bacterial lipoteichoic acids: an insight on its bio-
cytokine storm. Curr Opin Hematol 18:186–190.
logical significance. Org Biomol Chem 9:2040–2052.
Mima T, Nishimoto N. 2009. Clinical value of blocking IL-6 re-
Seifert H. 2009. The clinical importance of microbiological find-
ceptor. Curr Opin Rheumatol 21:224–230.
ings in the diagnosis and management of bloodstream infec-
Remick DG. 2007. Pathophysiology of sepsis. Am J Pathol
tions. Clin Infect Dis 48(Suppl 4):S238–S245.
Stearns-Kurosawa DJ, Osuchowski MF, Valentine C, Kurosawa
Remick DG, Bolgos G, Copeland S, Siddiqui J. 2005. Role of
S, Remick DG. 2011. The pathogenesis of sepsis. Annu Rev
interleukin-6 in mortality from and physiologic response to
sepsis. Infect Immun 73:2751–2757.
Suntharalingam G, Perry MR, Ward S, Brett SJ, Castello-Cortes
Remick DG, Bolgos GR, Siddiqui J, Shin J, Nemzek JA. 2002. Six
A, Brunner MD, Panoskaltsis N. 2006. Cytokine storm in a
at six: interleukin-6 measured 6 h after the initiation of sepsis
phase 1 trial of the anti-CD28 monoclonal antibody TGN1412.
predicts mortality over 3 days. Shock 17:463–467.
Riedemann NC, Guo RF, Ward PA. 2003. The enigma of sepsis. J
Tschaikowsky K, Hedwig-Geissing M, Braun GG, Radespiel-
Troeger M. 2011. Predictive value of procalcitonin, interleukin-
Rittirsch D, Hoesel LM, Ward PA. 2007. The disconnect between
6, and C-reactive protein for survival in postoperative patients
animal models of sepsis and human sepsis. J Leukoc Biol
with severe sepsis. J Crit Care 26:54–64.
Waage A, Brandtzaeg P, Halstensen A, Kierulf P, Espevik T.
Romano M, Sironi M, Toniatti C, Polentarutti N, Fruscella P,
1989. The complex pattern of cytokines in serum from patients
Ghezzi P, et al. 1997. Role of IL-6 and its soluble receptor in
with meningococcal septic shock. Association between inter-
induction of chemokines and leukocyte recruitment. Im-
leukin 6, interleukin 1, and fatal outcome. J Exp Med 169:333–338.
Rose-John S, Scheller J, Elson G, Jones SA. 2006. Interleukin-6
biology is coordinated by membrane-bound and soluble re-
ceptors: role in inflammation and cancer. J Leukoc Biol
Rose-John S, Waetzig GH, Scheller J, Grotzinger J, Seegert D.
2007. The IL-6/sIL-6R complex as a novel target for thera-
peutic approaches. Expert Opin Ther Targets 11:613–624.
Scheller J, Chalaris A, Schmidt-Arras D, Rose-John S. 2011. The
pro- and anti-inflammatory properties of the cytokineinterleukin-6. Biochim Biophys Acta 1813:878–888.
Received 9 July 2011/Accepted 9 September 2011
AU1: Headings are usually omitted at the beginning of an article. After the introductory paragraphs about
definitions of sepsis, can ‘‘Signal Transduction’’ be inserted as a heading?
AU2: Please use English transliteration of Greek word rather than symbols. AU3: Please provide an issue number for each journal reference. AU4: Is the Barkhausen reference in press? Couldn’t find this one on PubMed.
Estado de Santa Catarina PREFEITURA MUNICIPAL DE ÁGUAS MORNAS CNPJ: 82.892.266/0001-50 SECRETARIA DE ADMINISTRAÇÃO E FINANÇAS COMISSÃO PERMANENTE DE LICITAÇÃO PROCESSO N.º 18/2012 MODALIDADE: PREGÃO PRESENCIAL – SISTEMA DE REGISTRO DE PREÇOS TIPO DE LICITAÇÃO : MENOR PREÇO POR ITEM SESSÃO PÚBLICA PARA RECEBIMENTO DOCUMENTAÇÕES E RECEBIMENTO E ABERTURA DAS PROPO
The Carriwell Maternity Support Band has been tested by leading midwife Sister Lilian and has been found to dramatically reduce ligament pain, back ache and abdominal pressure during pregnancy. An expectant mom’s mid-section is extremely prone to aches and pains during pregnancy, and these are as many and varied in nature as the woman herself. The reasons for this discomfort include incr
 might be especially prone—is associated with dramaticallyincreased serum concentrations of pro-inflammatory cyto-kines, commonly known as ‘‘cytokine storm.’’ Although LPSsare intrinsic components of the outer membrane of gram-negative bacteria, lipoteichoic acids and peptidoglycans servea similar role in gram-positive sepsis (Schmidt and others2011).
might be especially prone—is associated with dramaticallyincreased serum concentrations of pro-inflammatory cyto-kines, commonly known as ‘‘cytokine storm.’’ Although LPSsare intrinsic components of the outer membrane of gram-negative bacteria, lipoteichoic acids and peptidoglycans servea similar role in gram-positive sepsis (Schmidt and others2011).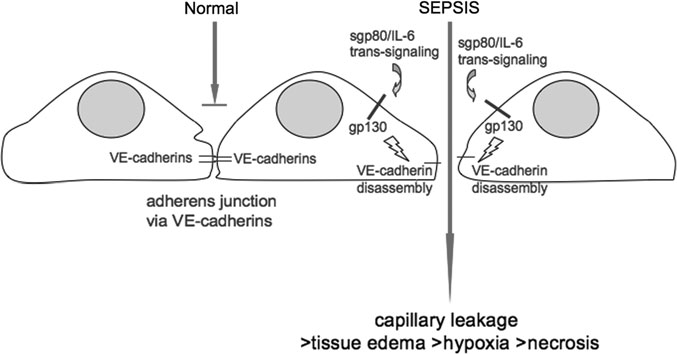 interleukin-6 (IL-6)/soluble IL-6R (sIL-6R) trans-signaling on vascular endo-thelial
Normally, adherens junctions betweenneighboring endothelial cells do notpermit uncontrolled influx of fluid fromthe blood into tissues. During sepsis,IL-6/sIL-6R trans-signaling triggersthe disassembly of these adherensjunctions by inducing the phosphory-lation of VE-cadherin. Influx of bloodinto tissues leads to edema, therebycontributing
lead to a drop in blood pressure with ensuing necrosis. This
Numerous explanations have been brought forward for
is a very serious consequence of shock-induced ischemia: The
resulting necrosis of the gut–blood barrier leads to an irre-
First, interfering with a complex system is complex
sistible invasion of the countless gut-resident commensal
bacteria, to which resistance is futile. Interestingly, Bar-
Second, lack of appropriate stratification of patients might
khausen and others (2011) recently showed in a sepsis model
have played a role in disappointing statistical effects; sepsis
that blocking IL-6 trans-signaling using sgp130Fc blocked
encompasses many different entities that have different
intestinal epithelial cell apoptosis in the gut and decreased
prognoses and different therapeutic needs at different time
mortality. Global blockade of IL-6 by a monoclonal antibody
points (Bone 1995). Support for ‘‘personalized sepsis medi-
had no beneficial effect. Thus, by keeping enteric epithelial
cine’’ is based on the finding that blunting inflammation
cells alive, blocking IL-6 trans-signaling protects patients
improves survival only of animals at a high risk of dying,
with sepsis by keeping the important barrier to enteric bac-
whereas low-dose glucocorticoids are effective in patients
teria sealed, protecting from a ‘‘second front’’ against those
with adrenal malfunction (Remick 2007). The need for a
opportunistic pathogens that threaten to invade the hosts’
tailored therapy in sepsis is already indicated by the obser-
vation that IL-6 levels are significantly higher in gram-negative bacteremia than in gram-positive bacteremia
(Alexandraki and Palacio 2010). Thus, if IL-6 indeed turnsout to be a valid therapeutic target in bacterial sepsis, the
Studies guided by a detailed understanding of the structure–
microbiological origin of sepsis surely needs to be taken into
function relationships of IL-6 with its receptors led to the
account, most likely resulting in differential dosing of IL-6
development of potent IL-6 inhibitors, such as monoclonal
antagonists according to the Gram state of the invading
neutralizing antibodies against IL-6 and its gp80 receptor, as
well as a soluble gp130 Fc fusion protein that inhibits IL-6/sIL-
Third, too late is too late: As a result of therapeutic in-
6R trans-signaling (Brakenhoff and others 1990; Brakenhoff
terference with first-line cytokines during the initial phase of
and others 1994; Grotzinger and others 1997). Previous studies
pathogenesis, therapy may have no benefit when initiated
that applied IL-6 inhibitors in murine disease models of ar-
too late (e.g., during the subsequent late phases of the clinical
thritis and colitis were encouraging and paved the way to
course when patients enter the hospital). Indeed, prospective
clinical studies (Rose-John and others 2007). As a result of such
studies on patients presenting with sepsis showed that in
persistent efforts, tocilizumab, a humanized monoclonal anti-
most patients, peak levels of key cytokines were present on
body that acts as an IL-6 receptor antagonist, has been ap-
the day of admission and decreased on the following days
proved for the treatment of rheumatoid arthritis (Mima and
(Damas and others 1997). Thus, the best time point for in-
terfering with first-line cytokines, such as IL-1b and TNF-a,
Although this seems to be good news for clinicians hoping
might have been missed. Encouragingly, IL-6 levels remain
that IL-6 antagonists will also be approved for the treatment
high and stable during the course of sepsis (Damas and
of sepsis, caution is warranted. So far, the history of the
others 1997). The study by Barkhausen and others (2011)
search for drugs to treat sepsis is so discouraging that this
demonstrated that a therapeutic application of the sgp130Fc
field has even been called the ‘‘pharmaceutical graveyard’’
protein 24 hours after the onset of the sepsis model still
(Riedemann and others 2003). Many of these failed endeav-
ors were aimed at interfering with cytokines known to exert
Fourth, there is a lack of appropriate animal models; mice
detrimental effects in sepsis. Why did so many previous
are mice, and humans are humans. As a species, humans are
expeditions—based on good basic science—eventually fail?
known to be particularly susceptible to lethal effects LPS.
interleukin-6 (IL-6)/soluble IL-6R (sIL-6R) trans-signaling on vascular endo-thelial
Normally, adherens junctions betweenneighboring endothelial cells do notpermit uncontrolled influx of fluid fromthe blood into tissues. During sepsis,IL-6/sIL-6R trans-signaling triggersthe disassembly of these adherensjunctions by inducing the phosphory-lation of VE-cadherin. Influx of bloodinto tissues leads to edema, therebycontributing
lead to a drop in blood pressure with ensuing necrosis. This
Numerous explanations have been brought forward for
is a very serious consequence of shock-induced ischemia: The
resulting necrosis of the gut–blood barrier leads to an irre-
First, interfering with a complex system is complex
sistible invasion of the countless gut-resident commensal
bacteria, to which resistance is futile. Interestingly, Bar-
Second, lack of appropriate stratification of patients might
khausen and others (2011) recently showed in a sepsis model
have played a role in disappointing statistical effects; sepsis
that blocking IL-6 trans-signaling using sgp130Fc blocked
encompasses many different entities that have different
intestinal epithelial cell apoptosis in the gut and decreased
prognoses and different therapeutic needs at different time
mortality. Global blockade of IL-6 by a monoclonal antibody
points (Bone 1995). Support for ‘‘personalized sepsis medi-
had no beneficial effect. Thus, by keeping enteric epithelial
cine’’ is based on the finding that blunting inflammation
cells alive, blocking IL-6 trans-signaling protects patients
improves survival only of animals at a high risk of dying,
with sepsis by keeping the important barrier to enteric bac-
whereas low-dose glucocorticoids are effective in patients
teria sealed, protecting from a ‘‘second front’’ against those
with adrenal malfunction (Remick 2007). The need for a
opportunistic pathogens that threaten to invade the hosts’
tailored therapy in sepsis is already indicated by the obser-
vation that IL-6 levels are significantly higher in gram-negative bacteremia than in gram-positive bacteremia
(Alexandraki and Palacio 2010). Thus, if IL-6 indeed turnsout to be a valid therapeutic target in bacterial sepsis, the
Studies guided by a detailed understanding of the structure–
microbiological origin of sepsis surely needs to be taken into
function relationships of IL-6 with its receptors led to the
account, most likely resulting in differential dosing of IL-6
development of potent IL-6 inhibitors, such as monoclonal
antagonists according to the Gram state of the invading
neutralizing antibodies against IL-6 and its gp80 receptor, as
well as a soluble gp130 Fc fusion protein that inhibits IL-6/sIL-
Third, too late is too late: As a result of therapeutic in-
6R trans-signaling (Brakenhoff and others 1990; Brakenhoff
terference with first-line cytokines during the initial phase of
and others 1994; Grotzinger and others 1997). Previous studies
pathogenesis, therapy may have no benefit when initiated
that applied IL-6 inhibitors in murine disease models of ar-
too late (e.g., during the subsequent late phases of the clinical
thritis and colitis were encouraging and paved the way to
course when patients enter the hospital). Indeed, prospective
clinical studies (Rose-John and others 2007). As a result of such
studies on patients presenting with sepsis showed that in
persistent efforts, tocilizumab, a humanized monoclonal anti-
most patients, peak levels of key cytokines were present on
body that acts as an IL-6 receptor antagonist, has been ap-
the day of admission and decreased on the following days
proved for the treatment of rheumatoid arthritis (Mima and
(Damas and others 1997). Thus, the best time point for in-
terfering with first-line cytokines, such as IL-1b and TNF-a,
Although this seems to be good news for clinicians hoping
might have been missed. Encouragingly, IL-6 levels remain
that IL-6 antagonists will also be approved for the treatment
high and stable during the course of sepsis (Damas and
of sepsis, caution is warranted. So far, the history of the
others 1997). The study by Barkhausen and others (2011)
search for drugs to treat sepsis is so discouraging that this
demonstrated that a therapeutic application of the sgp130Fc
field has even been called the ‘‘pharmaceutical graveyard’’
protein 24 hours after the onset of the sepsis model still
(Riedemann and others 2003). Many of these failed endeav-
ors were aimed at interfering with cytokines known to exert
Fourth, there is a lack of appropriate animal models; mice
detrimental effects in sepsis. Why did so many previous
are mice, and humans are humans. As a species, humans are
expeditions—based on good basic science—eventually fail?
known to be particularly susceptible to lethal effects LPS.