Tadalafil appartiene alla classe degli inibitori selettivi della fosfodiesterasi di tipo 5, con un profilo farmacocinetico caratterizzato da un’emivita terminale di circa diciotto ore. Dopo somministrazione orale viene assorbito rapidamente e raggiunge concentrazioni plasmatiche massime in due ore. La biotrasformazione avviene principalmente tramite CYP3A4 con formazione di metaboliti inattivi, escreti in prevalenza con le feci. L’elevato legame alle proteine plasmatiche (>90%) assicura una distribuzione stabile. Nei confronti delle altre molecole della stessa classe, cialis compresse italia è noto per la durata prolungata dell’attività farmacologica.
15krulwich.pm
J. Mol. Microbiol. Biotechnol. (2001) 3(2): 237-246.
Multifunctionality of Tetracycline Efflux Functions 237
JMMB Symposium Functions of Tetracycline Efflux Proteins that Do Not Involve Tetracycline Terry A. Krulwich*, Jie Jin, Arthur A. Guffanti, and David
transmembrane segments (TMS) and are encoded in the
H. Bechhofer
chromosome or on plasmids found predominantly in Gram-positive prokaryotes (Speer et al., 1992; Roberts, 1996;
Department of Biochemistry and Molecular Biology, Mount
McMurry and Levy, 2000). The Group 1 Tet proteins, by
Sinai School of Medicine, 1 Gustave L. Levy Place, New
contrast, are predominantly found in Gram-negative
prokaryotes and have 12-TMS. Both the 12-TMS and 14-TMS Tet proteins have significant deduced similarity toprokaryotic MDRs of the secondary antiporter type, e.g. Abstract
Bmr and Emr, respectively. Like such MDRs, the Tet effluxproteins are members of the Major Facilitator Superfamily
Tet(L) and Tet(K) are specific antibiotic-resistance
(MFS) of transport proteins (Marger and Saier, 1993). The
determinants. They catalyze efflux of a tetracycline(Tc)-
Tet proteins have been further classified together with
divalent metal complex in exchange for protons, as
MDRs and other drug efflux proteins in two families within
do other Tet efflux proteins. These Tet proteins also
the MFS, based on topological similarities rather than
catalyze Na+ and K+ exchange for protons. Each of the
clustering by specific substrate, i.e. “Drug:H + Antiporter
“cytoplasmic substrates”, Na+, K+ and the Tc-metal ion
(14-Spanner) or DHA14” and “Drug:H + Antiporter (12-
complex, can also be exchanged for K+, a catalytic
Spanner) or DHA 12” families (Paulsen et al., 1996; Pao
mode that accounts for the long-recognized K+ uptake
et al., 1998). Tet efflux proteins of both the DHA 12 and
capacity conferred by some Tet proteins. The multiple
DHA 14 families confer tetracycline (Tc)-resistance by virtue
catalytic modes of Tet(L) and Tet(K) provide potential
of their capacity to actively translocate a complex of Tc
new avenues for development of inhibitors of these
and a divalent metal ion that apparently bears a single net
efflux systems as well as avenues for exploration of
positive charge; Co2+ is generally the most effective in
structure-function relationships. The multiple catalytic
supporting transport, but Mg2+, Mn2+, Cd2+ and Ca2+ can
modes of Tet(L), which is chromosomally encoded in
also complex with Tc to form a substrate (Yamaguchi et
Bacillus subtilis, also correspond to diverse physiolog-
al., 1990; Yamaguchi et al., 1995; Guffanti and Krulwich,
ical roles, including roles in antibiotic-, Na+-, and alkali- resistance as well as K+ acquisition. The use of K+ as
Different groups and even classes of Tet efflux proteins
an external coupling ion may contribute not only to
exhibit different patterns with respect to the capacity to
the organism’s K+ uptake capacity but also to its ability
translocate diverse Tc analogues (Speer et al., 1992; Guay
to exclude Na+ and Tc at elevated pH values. Regulation
and Rothstein, 1993). Given this sort of “classical”
of the chromosomal tetL gene by Tc has been proposed
specificity and the absence of any known antibiotic
to involve a translational re-initiation mechanism that
substrates for Tet proteins other than Tc and its analogues,
is novel for an antibiotic-resistance gene and increases
the Tet efflux proteins cannot be described as bona fide
Tet expression seven-fold. Other elements of tetL
MDRs. Strategies for inactivating these problematic
expression and its regulation are already evident,
resistance determinants have accordingly focused on
including gene amplification and use of multiple
elements related to tet gene regulation and proliferation in
promoters. However, further studies are required to
response to exposure to Tc and on structure-based inhibitor
clarify the full panoply of regulatory mechanisms, and
development using Tc as the starting point (Nelson et al.,
their integration to ensure different levels of tetL
1993, 1994; Nelson and Levy, 1999; Schnappinger and
expression that are optimal for its different functions.
Hillen, 1996; Sum et al., 1998). But if not true MDRs, the
It will also be of interest to investigate the implications
Tet(L) and Tet(K) proteins are clearly multifunctional
of Tet(L) and Tet(K) multifunctionality on the
antiporters that are involved in resistance to multiple
emergence and persistence of these antibiotic-
stresses (Padan and Krulwich, 2000). These stresses,
resistance genes.
apart from antibiotic stress (Alekshun and Levy, 2000),include Na+ stress, alkali stress, and the challenge of K+
Introduction
insufficiency. Studies in our laboratory have shown thatTet(L) and Tet(K) utilize inorganic monovalent cations, Na+
Tet(L) and Tet(K)1, which are generally close to 60%identical to each other, are the major classes of tetracyclineefflux proteins (Tet proteins) among the Group 2 Tet proteins
1The nomenclature suggested by Levy et al. (1999) for tetracycline
(McMurry and Levy, 2000). These proteins have 14-
resistance determinants will be used in this article. Thus Tet(L) and Tet(K)will designate the proteins, and tetL and tetK will designate the genes ofclass L and K tetracycline resistance that were earlier named with indicationsof the allele, e.g. TetA(L) (Levy et al., 1989). For historical reasons (Williams
*For correspondence. Email [email protected];
and Smith, 1979), the B. subtilis tetL chromosomal locus was originally
Tel. (212) 241-7280; Fax. (212) 996-7214.
designated tetB and is referred to as such in the genome annotation.
Figure 1. Catalytic modes of Tet(L) and Tet(K).
and K+, as alternate cytoplasmic substrates to the Tc- -
surrounding the possibility that selective pressures other
Me2+ complex, i.e. Tet(L) and Tet(K) act as Na+(K+)/H+
than antibiotics may influence their retention and
antiporters (Cheng et al., 1994, 1996a,b,c; Guffanti and
Krulwich, 1995), even though divalent inorganic cationsare not transported except in complex with Tc (Yamaguchi
Catalytic Modes of Tet(L) and Tet(K)
et al., 1990; Cheng et al., 1996c). These proteins also havebeen shown to use K+ as well as H+ as the coupling ion
The three catalytic modes of Tet(L) and Tet(K) are depicted
whose entry down its electrochemical potential energizes
in Figure 1. Evidence for Mode I, the Tetracycline Efflux
the efflux of the cytoplasmic substrate (Guffanti et al., 1998).
mode, was anticipated by work with more intensively
These additional catalytic modes, which will be detailed
studied Tet proteins and was confirmed by Yamaguchi et
further below, have in common the use of cationic
al. (1995) and Guffanti and Krulwich (1995). Modes II and
substrates with single net charge, but the cytoplasmic
III, the Monovalent cation/H+ Antiport and Monovalent
substrates range from a Tc- -Me2+ complex that has regions
cation or Tc- -Me2+/K+ Antiport (the Net K+ Uptake Mode),
with highly non-polar features to highly hydrated
respectively, were unanticipated, although Ives and Bott
monovalent cations. The substrate diversity of Tet(L) and
(1990) speculated that tetL was likely to have a Tc-unrelated
Tet(K) opens up some of the same questions that are
function of physiological significance. Chromosomally
central to function of those MDRs whose substrates
encoded Tet(L) from Bacillus subtilis was identified as a
encompass diverse structures and both non-polar and polar
possible Na+/H+ antiporter after a mutant strain with a
compounds. The multiple catalytic modes of Tet(L) and
disruption in the tetL promoter region was isolated from
Tet(K), and their important physiological roles, also open
transposition libraries on the basis of Na+- and alkali-
up new approaches to structure-function analyses and to
sensitivity (Cheng et al., 1994; Krulwich et al., 1994). Na+/
development of Tet protein inhibitors. The multiple roles
H+ antiport activity, in addition to Tc- -Co2+ antiport, was
have several further implications. These include the
subsequently demonstrated in assays of everted
possible evolutionary relationship between these resistance
membrane vesicles of Escherichia coli strains expressing
determinants and house-keeping antiporters, and issues
either tetL or tetK (Cheng et al., 1994, 1996a; Guffanti and
Table 1. Cytoplasmic concentrations of Na+ after 15 h of growth of transformants of B. subtilis wild type and tetL deletion strain JC112 in malate mediumat pH 8.3 containing 100 mM Na+ and either 1 mM or 100 mM K+
Cytoplasmic Na+ (mM +/- standard deviation)
Data reproduced from Cheng et al., 1996b with permission.
Multifunctionality of Tetracycline Efflux Functions 239
Krulwich, 1995; Guffanti et al., 1998) and of
pACYC184, and Tet(K), in particular, had been reported to
proteoliposomes reconstituted with purified, C-terminally,
complement K+ uptake-deficient strains of E. coli (Dosch
hexa-histidine tagged Tet(L) (Cheng et al., 1996c). In the
et al., 1984; Griffith et al., 1988; Guay et al., 1993). It had
latter assays, 86Rb+ translocation in response to an
been suggested that the presence of one of these Tet
imposed pH gradient — an assay of K+/H+ antiport — was
proteins in the membrane, especially as expressed from a
also shown. Several observations indicated that the Tet-
multicopy plasmid, led to an electrogenic leak of K+ across
mediated Tc- -Me2+/H+ and Na+(K+)/H+ antiports (Modes I
the membrane (Guay et al., 1993), although Guay and
and II) are both electrogenic, i.e. the y:x ratio in Figure 1 is
Rothstein (1993) also noted the possibility that the capacity
greater than unity. If electrogenic in this manner, each
for K+ uptake could relate to the H+ or hydronium uptake
turnover results in a greater number of the coupling ions
associated with antiport energization. The N-terminal
entering a right-side-out system than the number of Na+,
domain of the Tet protein was particularly implicated in the
K+ or Tc- -Me2+ complexes effluxing. Net positive charge
promotion of K+ influx (Guay et al., 1993; Griffith et al.,
would thus be translocated inward during each turnover of
1988, 1994; Nakamura et al., 1995). Since Tet(K)
the transporter, accounting for the observation that abolition
possessed a strong K+/H+ antiport capacity, with H+/K+ >1,
of the transmembrane electrical potential (∆Ψ) markedly
we hypothesized that rather than a leak, the K+ uptake
inhibits these antiports (Guffanti and Krulwich, 1995;
mode might be a mode of the antiport (Mode III) in which
Guffanti et al., 1998) and, in proteoliposomes, an imposed
K+ serves as the external coupling ion (Guffanti et al., 1998).
transliposomal electrical potential can energize antiport
In the electrogenic exchange, even when K+ was also the
(Cheng et al., 1996c). The actual stoichiometry of Tet(L)
cytoplasmic substrate, net K+ uptake would result.
and Tet(K)-mediated antiports has yet to be determined.
Moreover, a controlled mode of the K+ antiport rather than
In the assays conducted thus far, both K+ and Li+ have
a K+ leak fit better with the observation that K+ uptake-
been found to inhibit Na+/H+ antiport but cross inhibition
deficient mutants of E. coli grew robustly when transformed
between Na+ and the Tc- -Me2+ complex has not yet been
with plasmids expressing tet(K). This would not be expected
demonstrated (Guffanti and Krulwich, 1995; Cheng et al.,
if the cost of K+ acquisition was an un-regulated
1996c). Lack of cross-inhibition might reflect the existence
electrogenic leak that would dissipate the ∆Ψ in an adverse
of distinct binding domains for the different substrates.
manner. Strong support for the Mode III, Net K+ Uptake
However, we would not yet rule out the possibility that the
Mode of the Tet antiport, was obtained in assays of right-
optimal assay conditions for demonstrating cross-inhibition
side-out vesicles from E. coli TK2420 (K+ uptake deficient)
have not yet been developed. This is an important issue to
expressing Tet(K) or Tet(L). An outwardly-directed gradient
resolve and is of particular interest given the report of
of Na+, K+, or Tc- -Me2+ drove electrogenic entry of 86Rb+;
multiple drug interaction sites in the lactococcal MDR, LmrP
the Rb+ accumulation was not supported by intravesicular
(Putman et al., 1999). Further definition of the pH profiles
choline, and not supported by Tet-less vesicles (Guffanti
of each substrate exclusion activity should also be of
et al., 1998; J. Jin, unpublished data). In a second
interest in delineating how the combination of modes works.
experimental protocol, in which intravesicular choline or
Initial findings indicate a greater Tc efflux activity at near
K+ and extravesicular K+(Rb+) were at equal
neutral pH and a greater Na+ efflux activity at elevated pH
concentrations, energy-dependent Rb+ accumulation did
not occur upon generation of the transmembrane ∆p in
It will also be of interest to quantify the relative Km
the choline-containing vesicles, as would have been
values of Tet(L) and Tet(K) for K+ and Na+ in the Na+(K+)/
predicted for a leak, but depended upon the presence of a
H+ antiport. A relatively higher K+ than Na+ preference for
Tet protein substrate inside the vesicles (Guffanti et al.,
Tet(K), compared to Tet(L), was indicated by the results of
1998). In both experimental protocols, the K+ uptake mode
a “Na + exclusion assay” from whole cells of tetL deletion
was more active in Tet(K) than in Tet(L) vesicles although
strain JC112 of B. subtilis (Cheng et al., 1996b). As shown
the activity of the latter was clear. It has not been shown
in Table 1, cells of wild type B. subtilis, with a normal
conclusively that K+ ions can serve as the sole coupling
chromosomal tetL and transformed with a control plasmid,
ions, rather than moving inward together with protons.
excluded 22Na+ effectively during growth in medium
However, recent data on the pH dependence of the Net K+
containing 100 mM Na+ and either 1 mM or 100 mM K+. In
Uptake Mode suggest that K+ can replace the H+ coupling
the presence of the higher K+ concentration, only a modest
elevation of Na+ accumulation in the cytoplasm was
Important questions are raised by the finding of the
observed relative to that found in the lower K+ medium ,
third, Net K+ Uptake Mode, apart from physiological
i.e. 43 vs. 32 mM. The tetL deletion strain, transformed
questions that will be considered below. Among these
with the control plasmid, exhibited greatly reduced capacity
questions are the following. First, does this mode extend
to exclude Na+. Upon transformation with a recombinant
the capacity of Tet(L) and Tet(K) proteins to support Tc
plasmid expressing tetL, the parental pattern was restored
and Na+ exclusion in the alkaline range of pH, driven by
to the mutant strain. By contrast, upon transformation with
the ∆Ψ, but independent of the reduced ∆pH (see Figure
a recombinant plasmid expressing tetK, effective Na+
1)? This question could be explored using recently isolated
exclusion was only observed in the low K+ medium.
mutants with a specific reduction in this Mode. Second, do
Once the Monovalent cation/H+ Antiport Mode (Figure
the original indications that expression of only the N-
1, Mode II) of both Tet(L) and Tet(K) had been established,
terminal one-quarter to half of Tet proteins can complement
we hypothesized that a long observed capacity of some
K+ uptake-deficient E. coli strains reflect a capacity of
Tet proteins to support K+ uptake might be another mode
truncated Tet proteins to catalyze some antiport modes?
of the Tet antiport activities. Tet(C), e.g. from pBR322 or
Can truncated forms catalyze electrogenic monovalent
Cytoplasm Membrane D1 L Periplasm
Figure 2. Topological model of Tet(L) showing motifs summarized by Paulsen et al. (1996). The model fits the deduced sequence of the chromosomally-encoded B. subtilis Tet(L) to the topological data obtained experimentally for Tet(K) (Ginn et al., 1997; Hirata et al., 1998).
cation/K+ exchange or are the truncated forms really just
Monovalent cation/H+ Antiport and the Net K+ Uptake
leaking K+? An initial set of experiments on one set of highly
Modes reflects the sharing of common structural features
truncated Tet(L) and Tet(K) forms did not indicate a capacity
for the cytoplasmic and coupling ions in different
for antiport (Guffanti et al., 1998), but the strength of the
conformations of the transporter. This has been indicated
earlier observations makes it worthwhile to undertake more
for the SMR protein EmrE (Yerushalmi and Schuldiner,
extensive studies of different truncated forms if examples
2000). Finally, it will be of interest to determine whether
of such forms can be identifed that are substantially
the robust K+ uptake supported by Tet(C) may also reflect
assembled into the membrane. Third, it will be of great
the capacity of this Group 1 Tet protein for antiport modes
interest to probe whether the apparently greater K+
II and III. The Tc- -Co2+/H+ antiport catalyzed by TetB, the
translocation by Tet(K) relative to Tet(L) in both the
Tn10-encoded Tet efflux protein, which is a 12-TMS Group
Table 2. Tc-, Na+- and alkali-related phenotypes of B. subtilis tetL mutant types typified by strains JC112 and JC112C
B. pH Homeostasis in Wild type and JC112 Transformants
Cytoplasmic pH, 10 min after pH 7.5 -> 8.5 shift
Data for parts A and B, shown with standard deviations, were reproduced with permission from the publisher from Wang et al., 2000 and Cheng et al., 1996b,respectively.
Multifunctionality of Tetracycline Efflux Functions 241
1 Tet protein like Tet(C), has been proposed to catalyze an
mutants that lack Na+ efflux capacity also lack Tc- -Me2+
electroneutral exchange (Yamaguchi et al., 1991a). If the
efflux capacity and vice versa. Given hypotheses about
exchanges catalyzed by the 12-TMS Tet proteins are
specific motifs, more targeted questions can also be posed.
indeed electroneutral, they would not be expected to be
For example, Ginn et al. (2000) recently reported on a
able to achieve net K+ uptake via a K+/K+ exchange, but
detailed analysis of the Motif C of Tet(K), using site-directed
could do so with a Tc- -Co2+/K+ exchange. The possibility
mutagenesis of individual residues and studying their
of an electrogenic K+/H+(K+) antiport capacity for Tet(C)
effects upon the Tetracycline Efflux Mode of transport. The
would also be worth examining directly since K+ uptake is
capacity for Net K+ Uptake was assayed in a growth assay
observed in the absence of antibiotic in pBR322 and
only, as a general assessment of transporter functionality
pACYC184 transformed cells (Dosch et al., 1984; Griffith
to supplement data from direct membrane incorporation
et al., 1988; Nakamura et al., 1995).
assays. Because Motif C has been proposed to beimportant in the coupling aspects of the DHA 14
Opportunities for Structure-Function Studies
mechanism, i.e. to be an “antiporter-associated motif”(Varela et al., 1995), we have recently undertaken studies
Motifs that are common to DHA 12 and/or DHA 14 type
of selected residues within this same motif, in Tet(L) and
transporters in general have been reviewed in detail by
Tet(K), but carried out quantitative vesicle assays of the
Paulsen et al. (1996). A topological model of the
Net K+ Uptake Mode. Thus far, these studies strongly
chromosomally encoded Tet(L) from B. subtilis, showing
support the conclusion that important determinants of the
the motif regions in this particular protein, is presented in
capacity for K+ uptake, and the greater capacity in Tet(K)
Figure 2; it was drawn using the experimental data obtained
vs. Tet(L), reside in Motif C in the region surrounding a
for Tet(K) topology by Ginn et al. (1997) and Hirata et al.
highly conserved proline, P156 (J. Jin, unpublished data).
(1998). Extensive studies of mutations across the whole
It will be of particular interest to determine whether single-
molecule of 12-TMS Tet(B), especially by Yamaguchi and
site mutants of Tet(L) that are Tet(K)-like with respect to
colleagues, have provided a wealth of information. This
the high activity of the K+ Uptake Mode also exhibit the
includes data about the dispensability of particular residues
greater K+:Na+ preference ascribed to Tet(K) relative to
for Tc translocation, indications of residues and regions
that may be involved in antibiotic substrate binding, and
The other arena in which the catalytic activities that
evidence for the involvement of particular TMS in forming
do not involve Tc may be useful is that of Tet protein
the perimeter of a water-filled channel (Iwaki et al., 2000;
inhibitors. There are indications that amiloride inhibits
Kimura-Someya et al., 2000; Kubo et al., 2000, and see
Tet(L)-mediated Na+ fluxes (Guffanti and Krulwich, 1995).
McMurry and Levy, 2000). Hirata et al. (1998), noting that
It will be of interest to more fully examine the Na+ and/or
Tet(B) and Tet(K) share at least some important residues
K+ related activities of these proteins as possible new
of comparable charge in analogous positions, suggested
avenues for inhibition of the transporter as a whole.
the possibility that the fundamental 3-dimensional structureof these 12- and 14-TMS Tet proteins will turn out to be
Physiological Roles and Regulation of 14-TMS Tet
similar, but direct structural information is not yet available. Proteins
Although specific information about regions and residuesin Tet(K) has begun to emerge with respect to the Tc- -
The physiological roles of B. subtilis Tet(L), that could also
Me2+ Efflux Mode (Fujihara et al., 1996; Ginn et al., 2000),
be supported by Tet(K), are shown in Figure 1. These roles
this information is still too fragmentary to assess the extent
were inferred from the phenotypes of mutants with an
of the correspondence between key residues and regions
insertion of a chloramphenicol-resistance cassette in place
of the 14- and 12-TMS Tet proteins. Nor has the oligomeric
of the tetL gene (Cheng et al., 1996b; Wang et al., 2000).
state of 14-TMS Tet proteins been established, a property
Ives and Bott (1990) had earlier been unable to isolate
that can be important for developing precise mechanistic
tetL deletion mutants, which led them to propose, as already
models. Studies of separate expression of two equal-sized
noted, that this locus probably had an important
domains from closely related 12-TMS Tet proteins had
physiological role. Perhaps sensitivities that were not yet
earlier shown apparent domain interactions that led to
anticipated, especially to [Na+], complicated there deletion
activity and that could involve intermolecular interactions
attempts. Even our successfully isolated mutant strains are
(Rubin and Levy, 1990, 1991). More recently, McMurry and
complicated by a pleiotropy in which two dominant types
Levy (1995) showed that the N-terminal half of Tet(B)
have been noted in a single isolation protocol (Cheng et
contains a “dimerization domain” and presented a general
al., 1996b; Wang et al., 2000). One type, which is less
model for intra and intermolecular interactions. Dimers and
frequently found and the more severely affected, is typified
higher oligomers of several secondary transporters from
by the mutant strain JC112. The more frequently found
eukaryotes have been proposed (e.g. Casey and
and less affected mutant type is typified by strain JC112C.
Reithmeier, 1991; Fafournoux et al., 1994; Kilic and
The Tc- and Na+-sensitivities of these mutants, and the
restoration of the wild type level of resistance by re-
As the structure-function relationships in 14-TMS Tet
introduction of a functional tetL gene, are shown in Table
proteins are further probed, there will be value added if
2A. JC112C exhibits wild type resistance to Na+, even at
the studies encompass effects on the other modes of
alkaline pH, and exhibits a Tc-sensitivity that is intermediate
transport, i.e. utilizing cytoplasmic substrates other than a
Tc- -Me2+ complex or using K+ instead of H+. At the simplest
The frequency with which tetL deletion is achieved,
level, it will be of interest to examine the extent to which
i.e. with no lower frequency than a comparable disruption
B.subtilisStrains
Figure 3. Dependence of the growth of wild type B. subtilis and two phenotypic types of tetL deletion mutants on the [K+] of the medium. The data arereplotted from Wang et al. (2000).
of the dispensable amyE locus, makes it unlikely that the
proteins that occurred as part of the putative suppression
mutational loss of tetL is rapidly lethal in B. subtilis.
However, there may be strong enough selective pressure
The control of tetB expression from Tn10, like the
to promote the appearance of suppressor mutations, as
genes encoding several other 12-TMS Tet proteins, is
has been observed in a multiply Na+/H+ antiporter-deficient
mediated by a repressor that is expressed from a divergent
strain of E. coli (Harel-Bronstein et al., 1995). We have
promoter (Bertrand et al., 1983). By contrast, plasmid and
hypothesized that there are different paths of suppression,
chromosomally encoded Tet(K) and Tet(L) proteins have
of differing efficacy, that may be adopted in such
been presumed to utilize largely, if not entirely, translational
circumstances (Wang et al., 2000). The transporter(s) that
attenuation control mechanisms because of the presence
accounts for the better “compensation” of the JC1 12C
of a leader sequence upstream of the structural gene
mutant type has not yet been identified. However,
(Hoshino et al., 1985; Khan and Novick, 1983; Mojumdar
expression of genes encoding two different transport
and Khan, 1988). Using transcriptional and translational
systems that are putatively involved with monovalent cation
fusions, increases in B. subtilis tetL expression up to 20-
homeostasis in B. subtilis were found to be elevated in the
fold have been found in response to Tc. Using diverse
tetL deletion mutants. This elevation was not reversed by
mutations in the leader region, we found that a significant
re-introduction of a functional tetL gene (Wang et al., 2000)
portion of this Tc-regulated expression was best accounted
and is not observed in mutants in another locus, e.g. amyE
for by a translational re-initiation model that had not earlier
locus, prepared in parallel. Clarification of the events that
been proposed for an antibiotic-resistance gene
occur upon tetL disruption in B. subtilis will be of interest
(Stasinopoulos et al., 1998). Preliminary data indicate that
as a potential avenue into the apparently complex systems
Tc also leads to a 3-fold increase in tetL mRNA stability by
physiology of this multifunctional gene.
a mechanism that has yet to be elucidated (S.
Using the JC112 mutant, in which Na+- and alkali-
Stasinopoulos, unpublished data). Mojumdar and Khan
sensitivity are evident, a role for Tet(L) in both Na+- and
(1988) had noted both an increase in tetK mRNA and a
K+-dependent pH homeostasis was shown in pH shift
higher increase in Tet protein upon induction of the tetK
experiments (Table 2B) (Cheng et al., 1996b). Tet(L) is
gene of pT181 by Tc in B. subtilis minicells. Additional
apparently physiologically important in cytoplasmic pH
elements that may, under some circumstances, contribute
regulation, a role that is supported by the poor growth of
to changes in Tet(L) levels were revealed by second-site
JC112 at alkaline pH (Cheng et al., 1996b). Both tetL mutant
mutations that restored wild type or higher levels of Tc-
strains also exhibit some growth deficit at pH 7.0 that may
resistance to B. subtilis strains that had been rendered
partly reflect a physiological role for the Net K+ Uptake
sensitive by deletion of the polynucleotide phosphorylase
Mode of Tet(L). As shown in Figure 3, the two mutant strains
gene. These mutations included: 4-5 fold tetL gene
are particularly deficient in their ability to grow at low [K+]
amplification; and a single nucleotide change in the tetL
(Wang et al., 2000). JC112, especially, exhibits some
promoter region that resulted in 10-fold higher transcription
residual growth deficiency at pH 7 even when a functional
from a normally low basal level (Bechhofer and
tetL is restored and no antibiotic or monovalent cation
Stasinopoulos, 1998). Two promoters for the monocistronic
stresses are imposed. This residual deficit may reflect a
tetL gene have been found in B. subtilis (Cheng et al.,
secondary effect of the elevation of multiple membrane
1996b; Bechhofer and Stasinopoulos, 1998). However,
Multifunctionality of Tetracycline Efflux Functions 243
Figure 4. Tetracycline uptake and accumulation and its anticipated reduction by either Na+/H+ antiport or the more specific and effective Tc– -Me2+/H+ antiport:part of the functional rationale for conjoining these activities.
transcriptional regulation that might account for the
particular have been proposed to have arisen on two or
observation of Na+- and alkali-sensitivity of the original
three separate occasions, presumably from other transport
transposition mutant, which has an insertion in the promoter
proteins (Sheridan and Chopra, 1991). The 12-TMS Tet
region (Cheng et al., 1994), has not yet been found. Nor is
and other DHA 12 proteins are thought to have arisen by
the basis for the low basal level of tetL expression
gene duplication from an ancestral 6-TMS transporter
understood. It is likely that the levels of tetL expression
(Levy, 1992; Paulsen and Skurray, 1993). Whereas the
that are needed to exclude Tc, which can partition across
14-TMS proteins such as Tet(L) and Tet(K) might have
the membrane (see section below), may be too high for
involved the fusion of the putative 6-TMS ancestor with an
optimal growth of B. subtilis under other conditions,
8-TMS interloper, features of the C-terminal 6-TMS of such
including the moderate stresses and challenges of Na+,
proteins have led to the alternate proposal that the DHA
alkali and K+ insufficiency in which Tet(L) function is
14 proteins resulted from the incorporation of two new,
involved. Different, more temperate mechanisms of control
central TMS in between the halves of a 12-TMS efflux
may obtain in response to these stresses, and it will be of
protein (Griffith et al., 1992; Paulsen and Skurray, 1993).
Perhaps the multifunctional Tet proteins, which may
include 12-TMS Tet(C) (Griffith et al., 1994) as well as the
Implications of 14-TMS Tet Multifunctionality for
14-TMS Tet(L) and Tet(K), evolved from structurally related
Theories of Origin and for Conferral of Competitive
transport proteins that have monovalent cation/H+ antiport
activity. The genome of B. subtilis contains numerous geneswhose products have strong sequence similarity and similar
The antibiotic-producing organisms are an obvious source
predicted topologies to Tet(L), as ascertained using BLAST
of “the ancestral” antibiotic resistance determinants. To
(Altschul et al., 1990) for example. The best matches
the extent that production of antibiotics serves an offensive
among dozens of such gene products lack specific residues
function in the ecosystem for the producer, these organisms
that are believed to be essential for Tc efflux, e.g. E152.
must effectively efflux the antibiotic (Saunders, 1984;
Perhaps one or more of these proteins nonetheless has
Cundliffe, 1989). However, sequence analyses of diverse
some modest Tc efflux capacity and/or have monovalent
types of multi-drug and specific drug efflux proteins (Saier
cation-related functions. We have hypothesized (Cheng
et al., 1998) suggested that the capacity for drug efflux
et al., 1996a) that there is an inherent “logic” to the
emerged in each category of membrane transport proteins
conjoining of this antiport capacity with that which excludes
from proteins that had their own physiological roles,
Tc. As illustrated in Figure 4, Tc is proposed to be
although this was a rare event. Subsequently, the primordial
predominantly charged at near neutral pH values but the
drug transporter was maintained over a long time period
uncharged component would partition across the
during which the substrate specificity underwent
membrane in a carrier-independent manner (Nikaido and
considerable change and diversification. Tet proteins in
Thanassi, 1993; Sigler et al., 2000). Yamaguchi et al.
(1991b) presented evidence that protonation of the
et al., 1986; Udo and Grubb, 1996). Simpson et al. (2000)
abundant Tc form that bears net negative charge results in
recently studied the competitive fitness of S. aureus that
carrier-independent, ∆pH-dependent Tc accumulation.
expressed tetK from plasmid pT181 vs. chromosomally
Even though it has been suggested that inward fluxes of
integrated forms of tetK that were expressed from different
both Tc forms are likely to be slow (Sigler et al., 2000), Tc
promoter configurations. They found that the
would accumulate more and be most toxic at pH values in
chromosomally integrated form, in spite of a lower gene
the neutral and acidic range, at which the ∆pH component
dosage, exhibited higher Tc resistance and higher
of the ∆p was maximal. Moreover, enhanced expression
competitive fitness than the S. aureus carrying pT181. The
of monovalent cation/H+ antiporters, e.g. the Nha shown
enhancement of competitive fitness was only observed in
in Figure 4, would reduce Tc toxicity even without any
the presence of Tc in these studies, but it would be of
capacity for Tc efflux since they would reduce the ∆pH. A
interest to re-examine the different constructs under
Tet protein that both lowered the ∆pH and catalyzed active
conditions that included Na+-, alkali- or low K+ challenge.
extrusion of the Tc- -Me2+ complex would, of course, be farmore effective for Tc exclusion. Having acquired the added
Acknowledgement
capacity to catalyze Tc- -Me2+/H+(K+) antiport, a Tet protein
The work in the authors’ laboratories was supported by research grant
that evolved from a Na+ and/or K+/H+(K+) antiporter might
GM52837 from the National Institute of General Medical Sciences.
at least sometimes retain all these catalytic capacities, as
References
observed in Tet(L) and Tet(K). Even if all the substrates ofsuch multifunctional antiporters turn out to utilize a common
Alekshun, M.N., and S.B. Levy. 2000. Bacterial drug resistance: response
cytoplasmic binding site on the Tet protein, different pH-
to survival threats. In: Bacterial Stress Responses. (G. Storz and R.
dependence for binding of the different substrates could
Hengge-Aronis, eds) pp. 323-366, ASM Press, Washington, D.C.
Altschul, S.F., W. Gish, W. Miller, E.W. Myers, and D.J. Lipman. 1990. Basic
prioritize the most physiologically important catalytic mode
local alignment search tool. J. Mol. Biol. 215: 403-410
under particular conditions of pH. The Tc exclusion activity
Amano, H. , C. L. Ives, K. F. Bott, and K. Shishido. 1991. A limited number
(Mode I) is most important in the neutral to acidic pH range
of Bacillus subtilis strains carry a tetracycline-resistance determinant at asite close to the origin of replication. Biochim. Biophys. Acta. 1088: 251-
where Tc is most toxic, whereas the Na+- and alkali-
resistance functions (Mode II and III) would be most
Bechhofer, D.H., and S.J. Stasinopoulos. 1998. tetA(L) mutants of a
important in the alkaline range (Padan and Krulwich, 2000).
tetracycline-sensitive strain of Bacillus subtilis with the polynucleotide
Might the multiple functions of Tet proteins also provide
phosphorylase gene deleted. J. Bacteriol. 180: 3470-3473
Bertrand, K.P., K. Postle, L.V. Wray, and W.S. Reznickoff. 1983. Overlapping
selective pressure for retention of tet genes independently
divergent promoters control expression of Tn10 tetracycline resistance.
of whether Tc was in abundance in the environment? This
is an important issue to consider in light of the findings
Bouma, J.E., and R.E. Lenski. 1988. Evolution of a bacteria/plasmid
with Tet(L) and Tet(K) and the current focus on reducing
Casey, J.R. and R.A. Reithmeier (1991) Analysis of the oligomeric state of
antibiotic levels in the environment as a means of reducing
Band 3, the anion transport protein of the human erythrocyte membrane,
the prevalence of resistance determinants (Levy, 1997).
by size exclusion high performance liquid chromatography. Oligomeric
In this connection, a finding of Lenski and colleagues
stability and origin of heterogeneity. J. Biol. Chem. 266: 15726-15737
Cheng, J., K. Baldwin, A.A. Guffanti, and T.A. Krulwich. 1996a. Na+/H+
(Bouma and Lenski, 1988; Lenski et al., 1994; Lenski, 1997)
antiport activity conferred by Bacillus subtilis tetA(L), a 5' truncation product
is intriguing. These investigators studied the competitive
of tetA(L), and related plasmid genes upon Escherichia coli. Antimicrob.
fitness of plasmid-bearing E. coli that had “co-evolved”
Cheng J., A.A. Guffanti, and T. A. Krulwich. 1994. The chromosomal
over 500 generations with pACYC184. The question posed
tetracycline resistance locus of Bacillus subtilis encodes a Na+/H+
was whether, after prolonged co-existence mandated by
antiporter that is physiologically important at elevated pH. J. Biol. Chem.
the presence of Tc or chloramphenicol, the separated, co-
evolved host or plasmid would show a different response
Cheng, J., A.A. Guffanti, W. Wang, T.A. Krulwich, and D.H. Bechhofer.
1996b. Chromosomal tetA(L) gene of Bacillus subtilis: regulation of
than naive host or plasmid in a new engagement. Normally
expression and physiology of a tetA(L) deletion strain. J. Bacteriol. 178:
a plasmid imposes a burden on a host in the absence of
antibiotic selection. The co-evolved plasmid exhibited no
Cheng, J., D.B. Hicks, and T. A. Krulwich. 1996c. The purified Bacillus subtilis
tetracycline efflux protein TetA(L) reconstitutes both tetracycline-cobalt/
change in the imposition of this burden when introduced
H+ and Na+/H+ exchange. Proc. Natl. Acad. Sci. USA. 93: 14446-14451
into a new “naive” host E. coli. By contrast, the co-evolved
Cundliffe, E. 1989. How antibiotic-producing organisms avoid suicide. Ann.
E. coli host that had been cured of the plasmid no longer
showed a burden when fresh, naive pACYC184 was
Dosch, D.C., F.F. Salvacion, and W. Epstein. 1984. Tetracycline element of
pBR322 mediates potassium transport. J. Bacteriol. 160: 1188-1190
reintroduced (Bouma and Lenski, 1988). In fact, a benefit
Fafournoux, P., J. Noel, and J. Pouyssegur. 1994. Evidence that the Na+/
was observed. Subsequent studies showed that this benefit
H+ exchanger isoforms NHE1 and NHE3 exist as stable dimers in
was specifically related to the tetC gene of the plasmid
membranes with a high degree of specificity for homodimers. J. Biol. Chem. 269: 2589-2596
(Lenski et al., 1994). Since tetC, in particular, introduces
Fujihira, E., T. Kimura, Y. Shiina, and A. Yamaguchi. 1996. Transmembrane
capacities for K+ uptake as well as Tc-resistance, the
glutamic acid residues play essential roles in the metal-tetracycline/H+
possibility is raised that this and the even more
antiporter of Staphylococcus aureus. FEBS Lett. 391: 243-246
multifunctional Tet(K) and Tet(L) proteins would offer
Gillespie, M.T., J.W. May, and R.A. Skurray. 1986. Detection of an integrated
tetracycline resistance plasmid in the chromosome of methicillin-resistant
antibiotic-unrelated advantages to even a modestly
Staphylococcus aureus. J. Gen. Microbiol. 132: 1723-1728
adapted host, depending upon the environmental
Ginn, S.L., M.H. Brown, and R.A. Skurray. 1997. Membrane topology of
conditions. There are tetL naive strains of B. subtilis that
the metal-tetracycline/H+ antiporter TetA(K) from Staphylococcus aureus. J. Bacteriol. 179: 3786-3789
could be used to examine this issue (Amano et al., 1991).
Ginn, S.L., M.H. Brown, and R.A. Skurray. 2000. The TetA(K) Tetracycline/
In this connection, it is notable that tetK has been found to
H+ antiporter from Staphylococcus aureus: mutagenesis and functional
incorporate into the chromosome of S. aureus (Gillespie
analysis of motif C. J. Bacteriol. 182: 1492-1498
Multifunctionality of Tetracycline Efflux Functions 245
Griffith, J.K., M.E. Baker, D.A. Rouch, M.G. Page, R.A. Skurray, I.T. Paulsen,
Levy, S.B., L.M. McMurry, V. Burdett, P. Courvalin, W. Hillen, M.C. Roberts,
K.F. Chater, S.A. Baldwin, and P.J. Henderson. 1992. Membrane transport
and D.E. Taylor. 1989. Nomenclature for tetracycline resistance
proteins: implications of sequence comparisons. Curr. Opin. Cell Biol. 4:
determinants. Antimicrob. Agents Chemother. 33: 1373-1374
Marger, M.D. and M.H. Saier, Jr. 1993. A major superfamily of
Griffith, J.K., D.H. Cuellar, C.A. Fordyce, K.C. Hutchings and A.A.
transmembrane facilitators that catalyse uniport, symport and antiport.
Mondragon. 1994. Structure and function of the class C tetracycline/H+
antiporter: three independent groups of phenotypes are conferred by
McMurry, L.M., and S.B. Levy. 1995. The NH2-terminal half of the Tn10
tetracycline efflux protein TetA contains a dimerization domain. J. Biol.
Griffith, J.K. T. Kogoma, D.L. Corvo, W.L. Anderson, and A.L. Kazim. 1988.
An N-terminal domain of the tetracycline efflux protein increases
McMurry, L.M, and S.B. Levy. 2000. Tetracycline resistance in Gram-positive
susceptibility to aminoglycosides and complements potassium uptake
bacteria. In: Gram Positive Pathogens (V.A. Fischetti, R.P. Novick, J.J.
defects in Escherichia coli. J. Bacteriol. 170: 598-604
Ferretti, D.A. Portnoy, J.I. Rood, eds.) pp 660-677, ASM Press,
Guay, G.G., and D.M. Rothstein. 1993. Expression of the tetK gene from
Staphylococcus aureus in Escherichia coli: comparison of substrate
Mojumdar, M., and S.A. Khan. 1988. Characterization of the tetracycline
specificities of TetA(B), TetA(C), and TetK efflux proteins. Antimicrob.
resistance gene of plasmid pT181 of Staphylococcus aureus. J. Bacteriol.
Guay, G.G., M. Tuckman, P. McNicholas, and D.M. Rothstein. 1993.
Nakamura, T., Y. Matsura, A. Ishihara, T. Kitagawa, F. Suzuki, and T.
Expression of the tet(K) gene from Staphylococcus aureus mediates the
Unemoto. 1995. N-terminal quarter part of tetracycline transporter from
transport of potassium in Escherichia coli. J. Bacteriol. 175: 4927-4929
pACYC184 complements K+ uptake activity in K+ uptake-deficient mutants
Guffanti, A.A., and T.A. Krulwich. 1995. Tetracycline/H+ antiport and Na+/
of Escherichia coli and Vibrio alginolyticus. Biol. Pharm. Bull. 18: 1189-
H+ antiport catalyzed by the Bacillus subtilis TetA(L) transporter expressed
in Escherichia coli. J. Bacteriol. 177: 4557-4561.
Nelson, M.L., and S.B. Levy. 1999. Reversal of tetracycline resistance
Guffanti, A.A., J. Cheng, and T.A. Krulwich. 1998. Electrogenic antiport
mediated by different bacterial tetracycline resistance determinants by
activities of the gram-positive Tet proteins include a Na+(K+)/K+ mode
an inhibitor of the Tet(B) antiport protein. Antimicrob. Agents Chemother.
that mediates net K+ uptake. J. Biol. Chem. 273: 26447-26454
Harel-Bronstein, M., P. Dibrov, Y. Olami, E. Pinner, S. Schuldiner, and E.
Nelson, M.L., B.H. Park, J.S. Andrew, V.A. Georgian, R.C. Thomas, and
Padan. 1995. MH1, a second- site revertant of an Escherichia coli mutant
S.B. Levy. 1993. Inhibition of the tetracycline efflux antiport protein by 13-
lacking Na+/H+ antiporters (∆nhaA∆nhaB), regains Na+ resistance and a
thio-substituted-5-hydroxy-6-deoxy-tetracycline. J. Med. Chem. 36: 370-
capacity to excrete Na+ in a ∆µH+-dependent fashion. J. Biol. Chem. 270:
Nelson, M.L., B.H. Park, and S.B. Levy. 1994. Molecular requirements for
Hirata, T., E. Fujihira, T. Kimura-Someya, and A. Yamaguchi. 1998.
the inhibition of the tetracycline antiport protein and the effect of potent
Membrane topology of the Staphylococcal tetracycline efflux protein Tet(K)
inhibitors on the growth of tetracycline-resistant bacteria. J. Med. chem.
determined by antibacterial resistance gene fusions. J. Biochem. 124:
Nikaido, H., and D.G. Thanassi. 1993. Penetration of lipophilic agents with
Hoshino, T., T. Ideda, N. Tomizuka, and K. Furukawa. 1985. Nucleotide
multiple protonation sites into bacterial cells: tetracyclines and
sequence of the tetracycline resistance gene of pTHT15, a thermophilic
fluoroquinolones as examples. Antimicrob. Agents Chemother. 37: 1393-
Bacillus plasmid: comparison with staphylococcal TcR controls. Gene 37:
Padan, E., and T.A. Krulwich. 2000. Sodium Stress. In: Bacterial Stress
Ives, C.L. and K.F. Bott. 1989. Cloned Bacillus subtilis chromosomal DNA
Responses (G. Storz and R Hengge-Aronis, eds) pp. 117-130, ASM Press,
mediates tetracycline resistance when present in multiple copies. J.
Pao, S.S., I.T. Paulsen, and M.H. Saier, Jr. 1998. Major Facilitator
Iwaki, S., N. Tamura, T. Kimura-Someya, S. Nada, and A. Yamaguchi. 2000.
Superfamily. Microbiol. Molec. Biol. Rev. 62: 1-34.
Cysteine-scanning mutagenesis of transmembrane segments 4 and 5 of
Paulsen, I.T., M.H. Brown, and R.A. Skurray. 1996. Proton-dependent
the Tn10-encoded metal-Tetracycline/H+ antiporter reveals a permeability
multidrug efflux systems. Microbiol. Rev. 60: 575-608
barrier in the middle of a transmembrane water-filled channel. J. Biol.
Paulsen, I.T. and R.A. Skurray. 1993. Topology, structure and evolution of
two families of proteins involved in antibiotic and antiseptic resistance in
Khan, S.A., and R.P. Novick. 1983. Complete nucleotide sequence of pT181,
eukaryotes and prokaryotes — an analysis. Gene 124: 1-11
a tetracycline-resistance plasmid from Staphylococcus aureus. Plasmid
Putman, M., L.A. Koole, H.W. van Veen, and W.N. Konings. 1999. The
secondary multidrug transporter LmrP contains multiple drug interaction
Kilic, F. and G. Rudnick. 2000. Oligomerization of the serotonin transporter
and its functional consequences. Proc. Natl. Acad. Sci. USA 97: 3106-
Roberts, M.C. 1996. Tetracycline resistance determinants: mechanisms of
action, regulation of expression genetic mobility, and distribution. FEMS
Kimura-Someya, T., S. Iwaki, S. Konishi, N. Tamura, Y. Kubo, and A.
Yamaguchi. 2000. Cycsteine-scanning mutagenesis around
Rubin, R.A., and S.B. Levy. 1990. Interdomain hybrid Tet proteins confer
transmembrane segments 1 and 11 and their flanking loop regions of
tetracycline resistance only when they are derived from closely related
Tn10-encoded metal-Tetracycline/H+ antiporter. J. Biol. Chem. 275: 18692-
members of the tet gene family. J. Bacteriol. 172: 2303-2312
Rubin, R.A., and S.B. Levy. 1991. Tet protein domains interact productively
Krulwich, T.A., J. Cheng, and A.A. Guffanti. 1994. The role of monovalent
to mediate tetracycline resistance when present on separate polypeptides.
cation/proton antiporters in Na+-resistance and pH homeostasis in Bacillus:
an alkaliphile versus a neutralophile. J. Exp. Biol. 196: 457-470
Saier, M.H., Jr., I.T. Paulsen, M.K. Sliwinski, S.S. Pao, R.A. Skurray, and H.
Kubo, Y., S. Konishi, T. Kawabe, S. Nada, and A. Yamaguchi. 2000. Proximity
Nikaido. 1998. Evolutionary origins of multidrug and drug-specific efflux
of periplasmic loops in the metal-Tetracycline/H+ antiporter of Escherichia
coli observed on site-directed cross-linking. J. Biol. Chem. 275: 5270-
Saunders, J.R. 1984. Genetics and evolution of antibiotic resistance. Brit.
Lenski, R.E. 1997. The cost of antibiotic resistance from the perspective of
Schnappinger, D. and Hillen, W. 1996. Tetracyclines: antibiotic action, uptake
the bacterium. In: Antibiotic resistance: origins, evolution, selection and
and resistance mechanisms. Arch. Microbiol. 165: 359-369
spread. Wiley, Chichester (Ciba Foundation Symposium 207) pp 131-
Sigler, A., P. Schubert, W. Hillen, and M. Niederweis. 2000. Permeation of
tetracyclines through membranes of liposomes and Escherichia coli. Eur.
Lenski, R.E., S.C. Simpson, T.T. Nguyen. 1994. Genetic analysis of a
plasmid-encoded host genotype- specific enhancement of bacterial fitness.
Simpson, A.E., R.A. Skurray, and N. Firth. 2000. An IS257-derived hybrid
promoter directs transcription of a tetA(K) tetracycline resistance gene in
Levy, S.B. 1992. Active efflux mechanisms for antimicrobial resistance.
the Staphylococcus aureus chromosomal mec region. J. Bacteriol. 182:
Antimicrob. Agents Chemother. 36: 695-703
Levy, S.B. 1997. Antibiotic resistance: an ecological imbalance. In: Antibiotic
Sheridan, R.P., and I. Chopra. 1991. Origin of tetracycline efflux proteins:
resistance: origins, evolution, selection and spread. Wiley, Chichester
conclusions from nucleotide sequence analysis. Mol. Microbiol. 5: 895-
Levy, S.B., L.M. McMurry, T.M. Barbosa, V. Burdett, P. Courvalin, W. Hillen,
Speer, B.S., N.B. Shoemaker, and A.A. Salyers. 1992. Bacterial resistance
M.C. Roberts, J.I. Rood, and D.E. Taylor. 1999. Nomenclature for new
to tetracycline: mechanisms, transfer, and clinical significance. Clin.
tetracycline resistance determinants. Antimicrob. Agents Chemother. 43:
Stasinopoulos, S.J., G.A. Farr, and D.H. Bechhofer. 1998. Bacillus subtilis
tetA(L) gene expression: evidence for regulation by translationalreinitiation. Mol. Microbiol. 30: 923-932
Sum, P.-E., F.-W. Sum, and S.J. Projan. 1998. Recent developments in
tetracycline antibiotics. Curr. Pharmaceut. Design 4: 119-132
Udo, E.E., and W.B. Grubb. 1996. A phage-mediated transfer of
chromosomally integrated tetracycline resistance plasmid inStaphylococcus aureus. Curr. Microbiol. 32: 286-290
Varela, M.F., C.E. Sansom, and J.K. Griffith. 1995. Mutational analysis and
molecular modelling of an amino acid sequence motif conserved inantiporters but not symporters in a transporter superfamily. Mol. Membr. Biol. 12: 313-319
Wang, W., A.A. Guffanti, Y. Wei, M. Ito, and T.A. Krulwich. 2000. Two types
of Bacillus subtilis tetA(L) deletion strains reveal the physiologicalimportance of TetA(L) in K+ acquisition as well as in Na+, alkali, andtetracycline resistance. J. Bacteriol. 182: 2088-2095
Williams, G., and I. Smith. 1979. Chromosomal mutations causing resistance
to tetracycline in Bacillus subtilis. Mol. Gen. Genet. 177: 23-29
Yamaguchi, A., Y. Iwasaki-Ohba, N. Ono, M. Kaneko-Ohdera, and T. Sawai.
1991a. Stoichiometry of metal-tetracycline/H+ antiport mediated bytransposon Tn10-encoded tetracycline resistance protein in Escherichiacoli. FEBS Lett. 282: 415-418
Yamaguchi, A., H. Ohmori, M. Kaneko-Ohdera, T. Nomura, and T. Sawai.
1991b. ∆pH-dependent accumulation of tetracyline in Escherichia coli. Antimicrob. Agents Chemother. 35: 53-56
Yamaguchi, A., Y. Shiina, E. Fujihira, T. Sawai, N. Noguchi, and M. Sasatsu.
1995. The tetracycline efflux protein encoded by the tet(K) gene fromStaphylococcus aureus is a metal-tetracycline/H+ antiporter. FEBS Lett. 365: 193-197
Yamaguchi, A., T. Udagawa, and T. Sawai. 1990. Transport of divalent
cations with tetracycline as mediated by the transposon Tn10-encodedtetracycline resistance protein. J. Biol. Chem. 265: 4809-4813
Yerushalmi, H. and S. Schuldiner. 2000. A common binding site for
substrates and protons in EmrE, an ion-coupled multidrug transporter. FEBS Lett. 476: 93-97.
Översikt över pågående kliniska prövningar för rivaroxaban Rivaroxaban är en ny s k direkt faktor Xa-hämmare i tablettform som prövas för en rad olika indikationer med målet att förebygga och behandla blodproppar. Rivaroxaban är den hittills bäst studerade direkta faktor Xa-hämmaren som befinner sig under utveckling. Över 20 000 patienter har redandeltagit i de genomförda
Technical Data Sheet TDS 1973 (IE) PROTIM® GREEN E Fungicidal and Insecticidal Green Wood Preservative FEATURES INSTRUCTIONS FOR USE Effective against wood boring insects For external applications, Protim Green E should be Protects against wood-rotting fungi applied during dry and if possible windless weather Suitable for use on all s
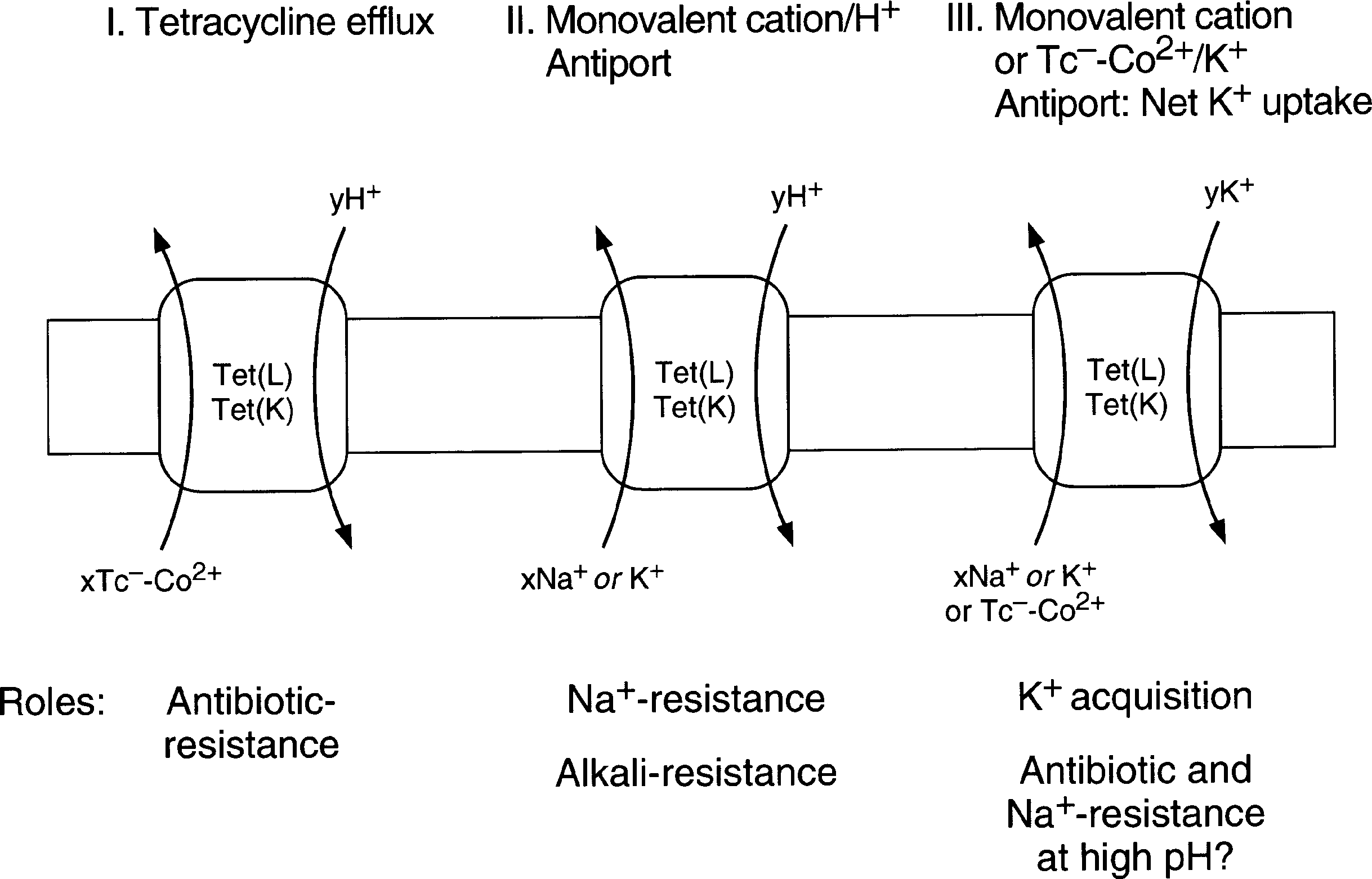 Figure 1. Catalytic modes of Tet(L) and Tet(K).
Figure 1. Catalytic modes of Tet(L) and Tet(K).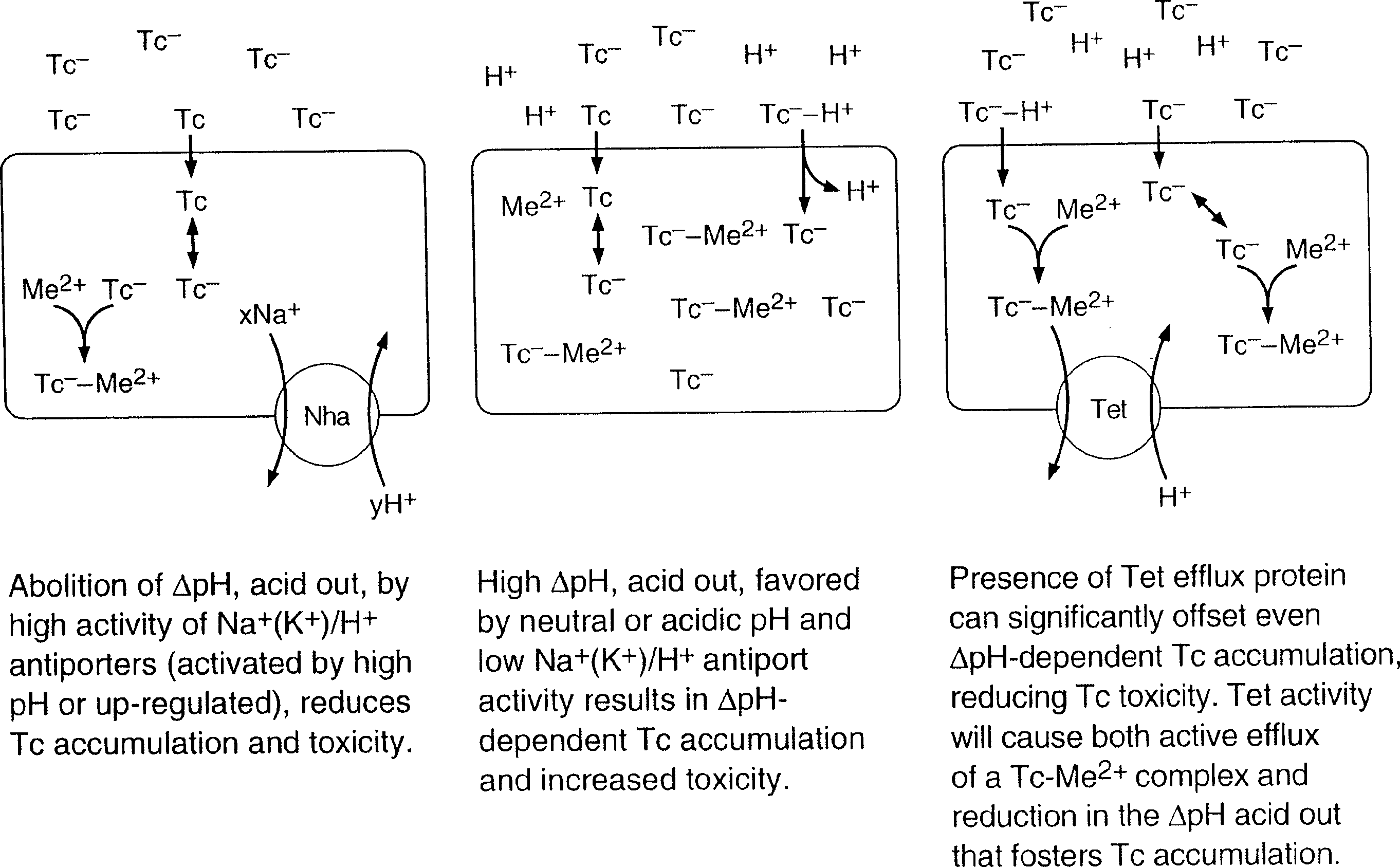 Multifunctionality of Tetracycline Efflux Functions 243
Figure 4. Tetracycline uptake and accumulation and its anticipated reduction by either Na+/H+ antiport or the more specific and effective Tc– -Me2+/H+ antiport:part of the functional rationale for conjoining these activities.
Multifunctionality of Tetracycline Efflux Functions 243
Figure 4. Tetracycline uptake and accumulation and its anticipated reduction by either Na+/H+ antiport or the more specific and effective Tc– -Me2+/H+ antiport:part of the functional rationale for conjoining these activities.