Tadalafil appartiene alla classe degli inibitori selettivi della fosfodiesterasi di tipo 5, con un profilo farmacocinetico caratterizzato da un’emivita terminale di circa diciotto ore. Dopo somministrazione orale viene assorbito rapidamente e raggiunge concentrazioni plasmatiche massime in due ore. La biotrasformazione avviene principalmente tramite CYP3A4 con formazione di metaboliti inattivi, escreti in prevalenza con le feci. L’elevato legame alle proteine plasmatiche (>90%) assicura una distribuzione stabile. Nei confronti delle altre molecole della stessa classe, cialis compresse italia è noto per la durata prolungata dell’attività farmacologica.
Doi:10.1016/j.ibmb.2006.05.009
Insect Biochemistry and Molecular Biology
Insect Biochemistry and Molecular Biology 36 (2006) 674–682
Xenobiotic response in Drosophila melanogaster: Sex dependence of
Gae¨lle Le Goffa,b,Ã, Fre´de´rique Hillioub, Blair D. Siegfriedc, Sam Boundya, Eric Wajnbergb,
Luc Soferb, Pascaline Audantb, Richard H. ffrench-Constanta, Rene´ Feyereisenb
aDepartment of Biology and Biochemistry, University of Bath, Bath BA2 7AY, UK
bINRA and Universite´ de Nice-Sophia Antipolis, UMR 1112, 06903 Sophia Antipolis, France
cDepartment of Entomology, University of Nebraska, Lincoln, Nebraska
Received 28 March 2006; received in revised form 19 May 2006; accepted 22 May 2006
The effect of xenobiotics (phenobarbital and atrazine) on the expression of Drosophila melanogaster CYP genes encoding cytochromes
P450, a gene family generally associated with detoxification, was analyzed by DNA microarray hybridization and verified by real-timeRT-PCR in adults of both sexes. Only a small subset of the 86 CYP genes was significantly induced by the xenobiotics. Eleven CYP genesand three glutathione S-transferases (GST) genes were significantly induced by phenobarbital, seven CYP and one GST gene wereinduced by atrazine. Cyp6d5, Cyp6w1, Cyp12d1 and the ecdysone-inducible Cyp6a2 were induced by both chemicals. The constitutiveexpression of several of the inducible genes (Cyp6a2, Cyp6a8, Cyp6d5, Cyp12d1) was higher in males than in females, and the inducedlevel similar in both sexes. Thus, the level of induction was consistently higher in females than in males. The female-specific andhormonally regulated yolk protein genes were significantly induced by phenobarbital in males and repressed by atrazine in females. Ourresults suggest that the numerous CYP genes of Drosophila respond selectively to xenobiotics, providing the fly with an adaptive responseto chemically adverse environments. The xenobiotic inducibility of some CYP genes previously associated with insecticide resistance inlaboratory-selected strains (Cyp6a2, Cyp6a8, Cyp12d1) suggests that deregulation of P450 gene expression may be a facile way to achieveresistance. Our study also suggests that xenobiotic-induced changes in P450 levels can affect insect fitness by interfering with hormonallyregulated networks. r 2006 Elsevier Ltd. All rights reserved.
Keywords: Phenobarbital; Atrazine; DNA microarray; CYP gene; Glutathione S-transferase; Esterase; Induction; Resistance
and 3-methylcholanthrene (Biochemicallydistinct P450 enzymes were revealed by these prototypical
Multigene families encoding cytochromes P450, glu-
inducers. Induction helped characterize most drug-meta-
tathione S-transferases and esterases are thought to
bolizing enzymes in vertebrates, before their total comple-
provide animals means to fend off xenobiotic challenges
ment was revealed by genome sequencing. In contrast, the
from the environment, such as toxic plant and microbial
large number of P450 genes in insects and their poor
chemicals encountered in their food or drugs, pesticides
representation in the initial EST collections (
and organic pollutants. The multiplicity of P450 enzymes
begged the question of their function. Few genes
was highlighted 40 years ago by the response of vertebrate
encoding insect P450s, glutathione S-transferases (GST) or
tissues to treatment by chemicals such as phenobarbital
esterases have been specifically and directly linked to a rolein detoxification or insecticide resistance despite the over-whelming biochemical evidence of their role in these
ÃCorresponding author. INRA and Universite´ de Nice-Sophia Anti-
polis, UMR 1112, 06903 Sophia Antipolis, France. Tel.: +33 4 92 38 65 78;
is therefore needed to characterize these genes. DNA
0965-1748/$ - see front matter r 2006 Elsevier Ltd. All rights reserved. doi:
G. Le Goff et al. / Insect Biochemistry and Molecular Biology 36 (2006) 674–682
microarray hybridization is the method of choice to study
genome-wide responses, as shown in Caenorhabditiselegans (). In Drosophila this
Total RNA was extracted from 50–100 flies using Trizol
technique has been used to identify responses to chemical
Reagent (Invitrogen Life technologies). Three independent
stressors such as paraquat, H2O2 and tunicamycin (
extractions were performed on three independent biologi-
microarrays have been used to detect genes that areconstitutively overexpressed in insecticide-resistant strains
from Drosophila and mosquitoes obtained from the field orselected further in the laboratory ;
A cDNA microarray was constructed using expressed
sequence tags (ESTs) of 1.8 kb average length and gene-
specific tags of 0.4 kb average length obtained by PCR
selectively represent all members of the detoxification gene
amplification and plasmid cloning. This ‘‘Drosophila
families and appropriate controls genes can provide a
toxicology microarray’’ contained 319 genes that represent
useful tool to identify inducible genes.
all cytochromes P450, glutathione S-transferases, esterases,
have used such a ‘‘detox chip’’ with 300 rat genes
other genes of interest including attacin or Ftz-F1 and
represented to study the effects of four known inducers,
several housekeeping genes as controls. Each insert was
phenobarbital, 3-methylcholanthrene, dexamethasone and
amplified by PCR with flanking universal primers, purified
clofibrate on rat liver gene expression. This study described
and then spotted using the ChipWriterPro Virtek (BioRad)
the inducer-specific pattern of transcriptional activation
spotter at three independent locations on Corning Ultra-
and confirmed the induction of CYP2B1, CYP2B2,
GAPS slides (Corning). All information concerning the
CYP2C6 as well as CYP3A12 and 3A2 by phenobarbital.
Induction of drug-metabolizing enzymes, and of cyto-
chrome P450-mediated pesticide metabolism in particular,has been extensively studied in insects ().
2.4. cDNA preparation and microarray hybridization
Induction of specific genes is now well documented (e.g.,CYP6A1; and induction of P450 genes
cDNA were synthesized from 10 mg of total RNA and
by a wide variety of chemicals is now amply demonstrated
labelled with the dyes Cy3- dCTP and Cy5- dCTP
(Amersham) using the Pronto! Universal Microarray
response within the whole multigene families or the
Hybridization Kit according to the manufacturer’s instruc-
selectivity of the inducer has yet to be documented.
tions. Each RNA was labelled with Cy3 and Cy5 to
Here we use a Drosophila melanogaster DNA microarray
perform reverse-labelled replicate arrays (dye swap) be-
of 319 genes covering in a redundant manner all CYP
cause of bias in dye incorporation. One comparison
genes, GST genes and esterase genes, to determine which
consists of a total of six arrays corresponding to two dye-
genes are affected in both males and females by treatment
swap arrays for each of our three biological replicates. We
of the flies with phenobarbital and atrazine, a prototypical
realized a balanced-block design of 24 arrays to compare
inducer and a widely used herbicide known to contaminate
phenobarbital and atrazine treatments in which female and
both terrestrial and aquatic habitats. Our results show that
male were separately treated. Hybridizations were per-
induction is sex- and inducer-specific and that xenobiotic
formed at 42 1C for 14 h, followed by post-hybridization
exposure can interfere with hormonally regulated physio-
washes. Slides were scanned on a Genepix 4000b scanner
and signal quantification was performed using the Genepixpro 4.1 software (Axon Instruments).
2.1. Drosophila stock and induction regime
Data files from Genepix were converted using Express
Converter software and normalized in Microarray Data
All fly stocks were maintained at 21 1C on standard corn
Analysis System (MIDAS), using two publicly available
medium for a 12:12 h dark/light period. Oregon R flies
were used as wild-type strain. Emerging flies were
Three steps of normalization were performed in MIDAS,
separated by sex and maintained on standard diet for
(a) a low-intensity filter in which signal intensity o1000
72 h before induction. The flies were then placed on
was removed; (b) a global lowess normalization was
medium containing the inducer at 10 mM for phenobarbi-
applied; (c) a flip dye consistency check, the Cy3 and Cy5
tal or 5 mM atrazine for 72 additional hours. These
labels were swapped between controls versus treatments to
concentrations were sublethal during the course of the
account for potential differences in labelling efficiency. For
experiment. After exposure, flies were frozen in liquid
statistical analysis, each array intensity was log-trans-
nitrogen for subsequent RNA extraction.
formed, centred by their average value and reduced by their
G. Le Goff et al. / Insect Biochemistry and Molecular Biology 36 (2006) 674–682
standard deviation. The resulting data for each gene were
treated versus control transcript levels expressed as fold
analyzed with a type III two-way (i.e., sex  treatment)
change for each treatment and for males and females.
analysis of variance (ANOVA) taking into account the
Phenobarbital induced seven P450 genes and one GST
presence of missing values. A gene was considered as
gene in males. In females, phenobarbital induced ten P450
differentially expressed if 50% or more of its variance was
and two GST genes and it down-regulated two P450 genes
explained by the treatment effect with a corresponding
and the a-Esterase 3 gene. Six P450 genes as well as GST
P-value lower than 0.035. Data were removed when there
D2 were induced by phenobarbital in both sexes. Atrazine
was no consistency for a same gene represented on the
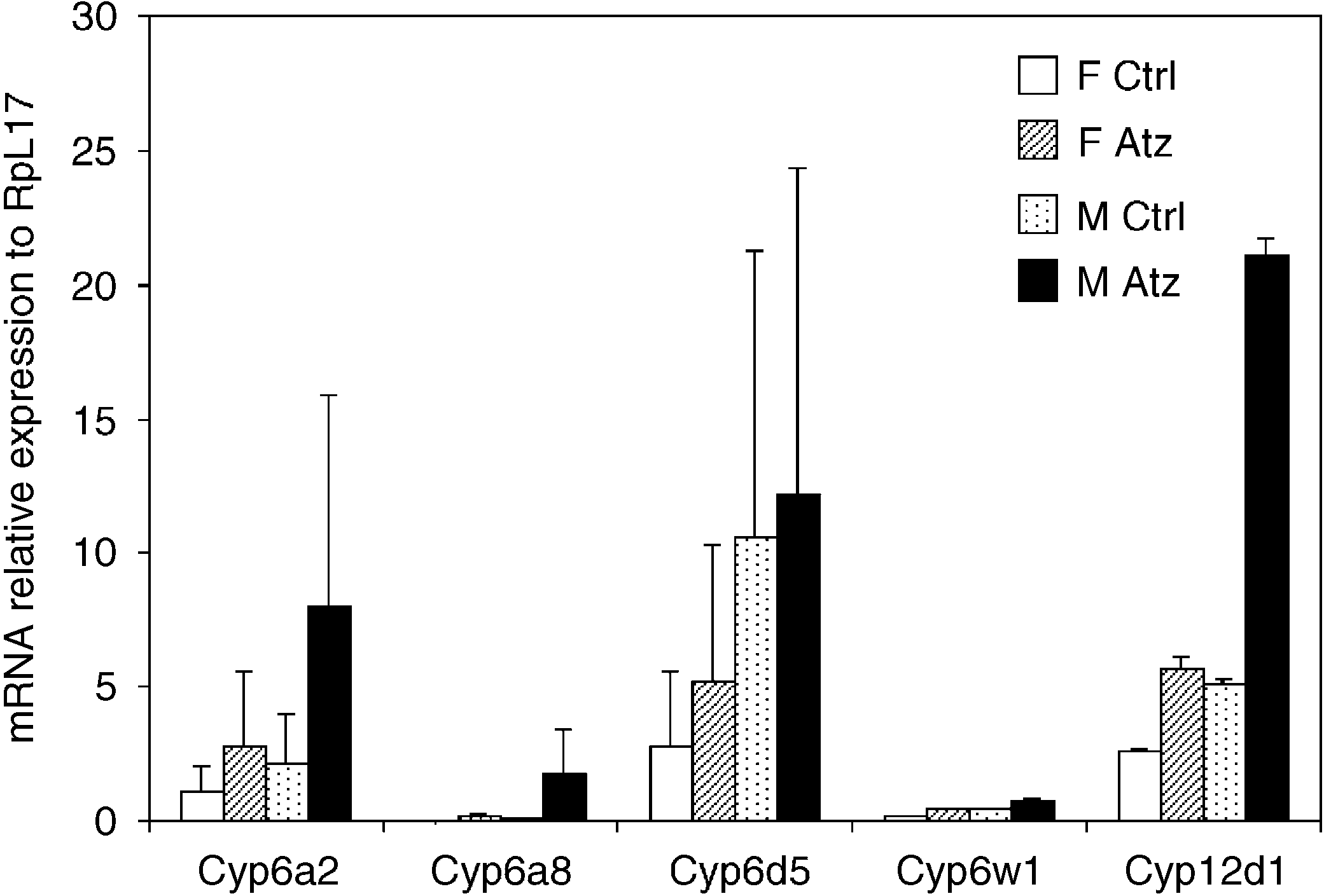
induced four P450 genes, a GST gene and an esterase gene
array by PCR products of different size, as this may have
in males; it repressed one P450 gene, a GST and an esterase
reflected a different degree of specificity.
gene. In females, atrazine induced six P450 genes. ThreeP450 genes were induced by atrazine in both sexes.
Only three P450 genes, Cyp6a2, Cyp6w1, Cyp12d1, were
induced by both chemicals and in both sexes. The other
Total RNA (1 mg) was reverse-transcribed using the
genes had sex- and/or chemical-selective responses. Sur-
iScript cDNA Synthesis Kit (BioRad). QPCR reactions
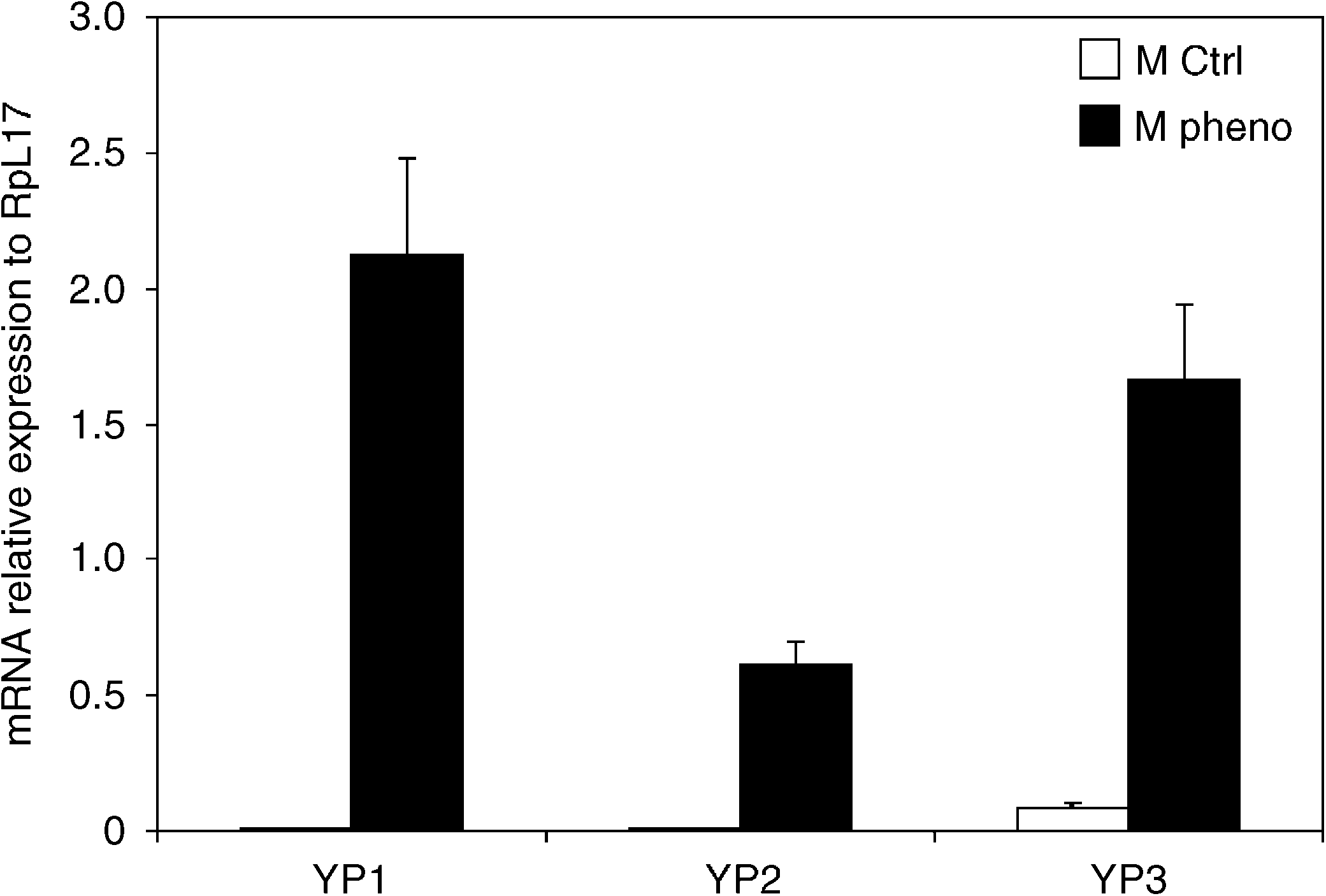
prisingly, the three yolk protein (Yp) genes that were part
were carried out on an Opticon monitor 2 (BioRad) using
of the set of control genes on the array were found to be
the Absolute QPCR SYBR green Mix (ABgene). The PCR
strongly induced by phenobarbital in males and were
conditions were as follows: 95 1C for 15 min to activate the
upregulated from virtually undetectable levels in untreated
hot-start DNA polymerase, followed by 40 cycles of 95 1C
flies. Atrazine down-regulated these same genes in females
for 30 s, 65 1C for 30 s and 72 1C for 30 s. Each reaction was
(). Further, the Ftz-f1 nuclear receptor gene was
performed in triplicate and the mean of three independent
significantly down-regulated in females by atrazine. Cyto-
biological replicates was calculated. All results were
chrome b5, a redox partner in some microsomal P450
normalized to the RpL17 mRNA level and calculated
reactions, was induced in males and repressed in females by
Microarray results were confirmed by qRT-PCR for
Cyp6a2, Cyp6d5, Cyp6g1, Cyp6w1, Cyp12d1 and also forCyp6a8 that was not included in the microarray experi-
Adult flies of both sexes were exposed to phenobarbital
ments for technical reasons. The results )
and atrazine in their food, and changes in the transcript
highlight the significant sex-dependent differences in
abundance for P450, GST and esterase genes assayed using
induction with low basal levels in females and more
microarray hybridization. list the genes that
spectacular induction than in males. The strong induction
were significantly up- or down-regulated and the ratio of
of yolk protein genes was confirmed by qRT-PCR ().
Table 1Genes differentially expressed in females flies after phenobarbital treatment (ANOVA)
G. Le Goff et al. / Insect Biochemistry and Molecular Biology 36 (2006) 674–682
Table 2Genes differentially expressed in male flies after phenobarbital treatment (ANOVA)
Table 3Genes differentially expressed in male flies after atrazine treatment (ANOVA)
Table 4Genes differentially expressed in female flies after atrazine treatment (ANOVA)
formly responsive to xenobiotics. Only five P450 genes, andone GST gene, were induced by the two chemicals used in
Our results show that the Drosophila genes coding for
this study, whereas another handful of genes showed a
the ‘‘drug-metabolizing enzymes’’ (DME) are not uni-
more selective pattern of transcriptional variation. Even
G. Le Goff et al. / Insect Biochemistry and Molecular Biology 36 (2006) 674–682
Fig. 3. Effects of phenobarbital on yolk protein mRNA levels in adultmales of Drosophila. Validations of microarray data by qPCR. Datapresented are the ratio of YP to Rpl17.
Fig. 1. Effects of phenobarbital on CYP mRNA levels in male and femaleDrosophila. Validations of microarray data by qPCR. Data presented are
a threshold of approximately 90% DNA identity (
whereas cDNA/PCR-based arrays candistinguish P450 genes up to 80% DNA identity ). We therefore used qRT-PCR normalizedagainst Rpl17 expression to confirm the results of themicroarrays. The results obtained are consistent with eachother, although qRT-PCR is, as expected, a more sensitivetool. The use of a microarray dedicated to a subset of geneshas already proven useful in detecting transcriptionalchanges that are associated with insecticide resistance inDrosophila ) andin the mosquito Anopheles gambiae ). Although whole genome arrays such as Affymetrix chipscan detect changes in more genes (e.g. ),practical considerations such as costs limit the number ofhybridizations and hence the statistical resolving power ofthese arrays. Thus, ‘‘detox chips’’ probably should becomea benchmark in the routine diagnosis of transcriptional
Fig. 2. Effects of atrazine on CYP mRNA expression in male and female
changes in the detoxification pathways of any organism
Drosophila. Validations of microarray data by qPCR. Data presented are
with a fully sequenced genome. Such thematic arrays have
already proven their utility in vertebrate toxicology() and plant physiology (
though our arrays cover by design only a small proportion
of all Drosophila genes, the results clearly identify a subset
Several of the genes shown to be transcriptionally
of genes that respond to xenobiotics and support the
regulated by phenobarbital or atrazine have been reported
usefulness of such thematic microarrays. Our study reveals
before to be associated with metabolic resistance to
that some of the genes regulated by xenobiotics have been
associated with insecticide resistance or shown to be
involved in resistance. Further we show that there is a
strong sex bias in the induction, and that treatment by
xenobiotics also disrupts the normal expression of genes
known to be involved in endocrine processes.
Some of these genes are also inducible by other chemicals,
The selective detection of transcriptional differences of
such as DDT (). Insecticide resistance is
closely related members of large gene families can be a
generally attributed to constitutive overexpression of these
difficult task, that was addressed for P450s in our study by
the use of multiple DNA features of different sizes for each
mutations in Cyp6a2 enable DDT metabolism. There are
gene on the array, as well as by additional techniques, such
other well-documented cases of xenobiotic-inducible genes
as qRT-PCR. Affymetrix chips can distinguish genes up to
that are constitutively overexpressed in insecticide-resistant
G. Le Goff et al. / Insect Biochemistry and Molecular Biology 36 (2006) 674–682
strains, particularly CYP6A1 and CYP6D1 in the house fly
(review in ). Is this relationship between
constitutive expression of a P450 gene was reported
induction and resistance coincidental or is there an
underlying mechanism that ought to be studied in greater
study shows that transcriptional regulation by xenobiotics
detail? Early work, predominantly on housefly P450, had
shown that resistant flies had biochemically distinct P450s,
Phenobarbital is one of the most commonly used
higher constitutive levels of P450, and different induction
inducers of cytochromes P450, known to be effective in
patterns. In some resistant strains induction is even greater
animals as well as plants, and even in bacteria (
). For insects, induction of a P450-dependent enzyme
resistant strains, P450 levels are similar to induced levels of
their susceptible counterparts (or even higher) and little or
no further induction can be observed ().
ago. Molecular studies in several insect species have now
Different inducers can also be distinguished in this way,
identified many phenobarbital-inducible genes (review in
some effective in a particular strain and others not
species, several P450 genes were inducible as CYP6A1,
CYP12A1 and CYP6D1 in the housefly, but here we
metabolic resistance and induction were associated with
provide a complete view of the pleiotropic effect of
the production of different forms of glutathione S-
phenobarbital induction on the CYP family in adult fruit
flies. Regulatory elements responsible for the phenobarbi-
induction in resistant flies was likely due to (P450) gene
tal response of the Cyp6a2 and Cyp6a8 genes of Drosophila
amplification, but they rejected induction as a factor in the
development of resistance in the field. It was suggested that
the same regulatory (trans-acting) gene may be involved in
phenobarbital-inducible genes should facilitate this mole-
both induction and biochemical resistance ),
and further proposed that a receptor involved
Atrazine is another known inducer of P450. Atrazine
in xenobiotic induction may be altered in resistant insects.
induces P450-dependent activities and a CYP4-related
However, despite growing evidence that overexpression of
P450 gene in the aquatic larvae of the midge Chironomus
some P450 genes in resistant strains can involve trans-
induction of P450 activities has also been reported in
induction has remained tenuous. Genetic analysis in
Drosophila indicates that expressions of Cyp6a2 and
induced pentoxyresorufin O-dealkylation activity in rats, as
Cyp6a8 are controlled in trans by a negative regulator
well as CYP2B on western blots, in a manner similar to
mutations alter the normal regulatory network involved in
Drosophila, we have shown that atrazine has a pattern of
the induction of a P450 gene, this could result in higher
induction that is similar, but not identical to phenobarbi-
constitutive expression of the gene and hence resistance.
tal, and that is characterized by the induction of several
The link between resistance and induction would thus be
GST genes in males. This indicates a degree of selectivity,
coincidental. Nonetheless, genes that are readily inducible
but also an underlying complexity of the induction
by xenobiotics have this regulatory mechanism in addition
response. A large-scale screen of gene expression in the
to their normal tissue, sex and developmental regulation.
nematode C. elegans revealed that CYP31A1 and A3 are
Thus, they offer a broader target for mutational events
inducible by both phenobarbital and atrazine whereas
(point mutations, duplications, transposable element inser-
CYP35A5 and 35C1 are induced by atrazine but not by
tions, etc.) and may be more likely to be involved in
resistance. Xenobiotic inducibility of a drug-metabolizing
more chemicals will uncover the xenobiotic regulation of
enzyme may therefore represent a risk factor for the
different genes, and that a picture will emerge of over-
lapping specificities of induction of P450 genes and/or
Some of the sex differences in gene expression may be
attributable to sex-linked differences in metabolism and
Closely related genes showed a differential response to
disposition of the inducers. In fact, little is known of
the two inducing compounds: for instance, Cyp6a19 was
phenobarbital or atrazine metabolism in insects (see
repressed by phenobarbital in females, but Cyp6a2, 9, 17
and Cyp6a23 were induced, whereas other CYP6 genes
exposure to the relatively high dose of inducer should
were not affected. Some genes may represent ‘‘stress-
minimize pharmacokinetic effects in favor of the pharma-
responsive genes’’ such as Cyp28a5. This gene was induced
by phenobarbital in both sexes, and by atrazine in males,
Sex-dependent gene expression in Drosophila is well
and this gene is also induced by paraquat, H2O2 and
function of the genes transcriptionally regulated by
G. Le Goff et al. / Insect Biochemistry and Molecular Biology 36 (2006) 674–682
phenobarbital or atrazine is largely unknown. Cyp6a2 is an
document whether xenobiotic treatments actually modify
abundant P450, enriched about 8-fold in Malpighian
hormone levels. Xanthotoxin inducibility of the CYP6B1
gene in the black swallowtail, Papilio polyxenes is
dependent on a xenobiotic response element in the gene
promoter. Drosophila transcription factors related to the
by many ESTs and the latter was first identified in olfactory
vertebrate aryl hydrocarbon receptor appear to modulate
organs (). Cyp12d1 is of special interest
expression of the CYP6B1 promoter, but a ligand-
because it encodes a mitochondrial P450, whereas the other
dependent factor is not yet been identified (
genes encode microsomal P450s. The related CYP12A1 of
The interactions between physiological signals and
the housefly is also inducible by phenobarbital, and
xenobiotics are necessary to allow the organism to
integrate information from its environment and adapt to
CYP12-like P450s of insects are thus a special type of
it. The role of the so-called drug-metabolizing enzymes in
inducible, environmental response genes, unlike the ster-
this interplay is becoming clearer (reviewed by
oid-metabolizing mitochondrial P450s of vertebrates and
). Our experiments start to shed some light on
the known effects of P450 inducers on insect fitness as
Little is known of the physiological substrates of the
phenobarbital treatment was shown to delay development
induced enzymes but Cyp6a2 was previously shown to be
and reduce fecundity in flies by a mechanism that has
metabolize JH III and related sesquiterpenoids (
more precisely the subset of xenobiotic-inducible genes
and Cyp18a1 was initially cloned as a result of
within the families encoding drug-metabolizing enzymes.
Tests on larvae as well as adults with a greater variety of
Lobster CYP45, a CYP6/9-like P450 is induced by
xenobiotics are thus warranted. The hypothesis that
phenobarbital and by 20-hydroxyecdysone (
inducibility is a risk factor for resistance as discussed
Similarly, CYP330A1 (a member of the CYP2 clan as
CYP18) is inducible by ecdysteroids and by phenobarbitalin the crab Carcinus maenas whereas CYP4C39 is notThese genes are thus possible cross-
over points between endocrine and xenobiotic signals. Interestingly, the bFTZ-F1nuclear receptor, down-regu-
We thank Charles Robin, Department of Genetics,
lated by atrazine in female flies, is known to regulate
University of Melbourne for clones of most of the GSTs
ecdysteroid titers in Drosophila by controlling the expres-
and esterase genes used on our array, Claire Conrad and
sion of the dib and phm genes (). The dib
Marcel Amichot for a set of P450 gene-specific tags, and
and phm genes (Cyp302a1 and Cyp306a1) are expressed at
Nathalie Tijet and Christian Helvig for their initial work
levels too low to be detected in our whole-body sampling
procedure. Nonetheless, the observation that yolk proteingene expression is down-regulated by atrazine as well asbFTZ-F1 is highly suggestive of endocrine disruption,
because ecdysteroids normally induce yolk protein synth-
Agosin, M., Scaramelli, N., Gil, L., Letelier, M.E., 1969. Some properties
esis in female flies. Endocrine disruption is also suggested
of the microsomal system metabolizing DDT in Triatoma infestans.
by the induction of yolk protein genes in males after
Comp. Biochem. Physiol. 29, 785–793.
phenobarbital treatment. Male flies do not normally
Amichot, M., Brun, A., Cuany, A., Helvig, C., Salaun, J.P., Durst, F.,
produce vitellogenin, and until now, only 20-hydroxyecdy-
Berge, J.B., 1994. Expression study of CYP genes in drosophila strains
sone excess was shown to induce yolk protein gene
resistant or sensitive to insecticides. In: Lechner, M.C. (Ed.),Cytochrome P450. John Libbey Eurotext, Paris, pp. 689–692.
Amichot, M., Tares, S., Brun-Barale, A., Arthaud, L., Bride, J.M., Berge,
). Xenobiotic interference with vitellogenesis has been
J.B., 2004. Point mutations associated with insecticide resistance in the
demonstrated before in invertebrates as the functional
Drosophila cytochrome P450 Cyp6a2 enable DDT metabolism. Eur. J.
homolog of yolk proteins, vitellogenin mRNA, is induced
by bisphenol-A and vinclozolin in C. elegans (
Andersen, J.F., Walding, J.K., Evans, P.H., Bowers, W.S., Feyereisen, R.,
1997. Substrate specificity for the epoxidation of terpenoids and active
site topology of house fly cytochrome P450 6A1. Chem. Res. Toxicol.
The exact mechanisms of these potential endocrine-
disrupting effects remain to be established. For instance,
Arbeitman, M.N., Fleming, A.A., Siegal, M.L., Null, B.H., Baker, B.S.,
we do not know yet whether the xenobiotics interact
2004. A genomic analysis of Drosophila somatic sexual differentiation
directly with hormone receptors or whether putative
and its regulation. Development 131, 2007–2021.
Bogwitz, M.R., Chung, H., Magoc, L., Rigby, S., Wong, W., O’Keefe, M.,
xenosensors such as DHR96, the Drosophila ortholog of
McKenzie, J.A., Batterham, P., Daborn, P.J., 2005. Cyp12a4 confers
PXR/CAR nuclear receptors of vertebrates, are the entry
lufenuron resistance in a natural population of Drosophila melanoga-
point of some xenobiotic signals. We need also to
ster. In: Proceedings of the National Academy of Science, USA.
G. Le Goff et al. / Insect Biochemistry and Molecular Biology 36 (2006) 674–682
Bownes, M., 1982. The role of 20-hydroxyecdysone in yolk-polypeptide
Hallstrom, I., Blanck, A., Atuma, S., 1984. Genetic variation in
synthesis by male and female fat bodies of Drosophila melanogaster.
cytochrome P-450 and xenobiotic metabolism in Drosophila melano-
gaster. Biochem. Pharmacol. 33, 13–20.
Brandt, A., Scharf, M., Pedra, J.H., Holmes, G., Dean, A., Kreitman, M.,
Hurban, P., Thummel, C.S., 1993. Isolation and characterization of fifteen
Pittendrigh, B.R., 2002. Differential expression and induction of two
ecdysone-inducible Drosophila genes reveal unexpected complexities in
Drosophila cytochrome P450 genes near the Rst(2)DDT locus. Insect
ecdysone regulation. Mol. Cell Biol. 13, 7101–7111.
Kao, L.M., Wilkinson, C.F., Brattsten, L.B., 1995. In vivo effects of 2,4-D
Brown, R.P., McDonnell, C.M., Berenbaum, M.R., Schuler, M.A., 2005.
and atrazine on cytochrome P-450 and insecticide toxicity in southern
Regulation of an insect cytochrome P450 monooxygenase gene
armyworm (Spodoptera eridania) larvae. Pestic. Sci. 45, 331–334.
(CYP6B1) by aryl hydrocarbon and xanthotoxin response cascades.
Kasai, S., Tomita, T., 2003. Male specific expression of a cytochrome P450
(Cyp312a1) in Drosophila melanogaster. Biochem. Biophys. Res.
Brun, A., Cuany, A., Le Mouel, T., Berge, J., Amichot, M., 1996.
Inducibility of the Drosophila melanogaster cytochrome P450 gene,
Kristensen, C., Morant, M., Olsen, C.E., Ekstrom, C.T., Galbraith, D.W.,
CYP6A2, by phenobarbital in insecticide susceptible or resistant
Lindberg Moller, B., Bak, S., 2005. Metabolic engineering of dhurrin
strains. Insect Biochem. Mol. Biol. 26, 697–703.
in transgenic Arabidopsis plants with marginal inadvertent effects on
Carin˜o, F., Koener, J.F., Plapp Jr., F.W., Feyereisen, R., 1992. Expression
the metabolome and transcriptome. Proc. Natl. Acad. Sci. USA 102,
of the cytochrome P450 gene CYP6A1 in the housefly, Musca
domestica. In: Mullin, C.A., Scott, J.G. (Eds.), Molecular Mechanisms
Kohra, S., Tominaga, N., Mitsui, Y., Takao, Y., Ishibashi, Y., Arizono,
of Insecticide Resistance. American Chemical Society, Washington,
K., 1999. Determination of a screening system of endocrine disruptors
by the induction of vitellogenin mRNA in C. elegans larvae. J. Health
Conney, A.H., 1967. Pharmacological implications of microsomal enzyme
induction. Pharmacol. Rev. 19, 317–366.
Le Goff, G., Boundy, S., Daborn, P.J., Yen, J.L., Sofer, L., Lind, R.,
Daborn, P.J., Yen, J.L., Bogwitz, M.R., Le Goff, G., Feil, E., Jeffers, S.,
Sabourault, C., Madi-Ravazzi, L., ffrench-Constant, R.H., 2003.
Tijet, N., Perry, T., Heckel, D., Batterham, P., Feyereisen, R., Wilson,
Microarray analysis of cytochrome P450 mediated insecticide resis-
T.G., ffrench-Constant, R.H., 2002. A single P450 allele associated
tance in Drosophila. Insect Biochem. Mol. Biol. 33, 701–708.
with insecticide resistance in Drosophila. Science 297, 2253–2256.
Liu, N., Scott, J.G., 1997. Phenobarbital induction of CYP6D1 is due to a
Darvas, B., Rees, H.H., Hoggard, N., Tag El-Din, M.H., Kuwano, E.,
trans acting factor on autosome 2 in house flies, Musca domestica.
Belai, I., Timar, T., 1992. Cytrochrome P-450 inducers and inhibitors
interfering with ecdysone 20-monooxygenases and their activities
Londono, D.K., Siegfried, B.D., Lydy, M.J., 2004. Atrazine induction of a
during postembryonic development of Neobellieria bullata Parker.
family 4 cytochrome P450 gene in Chironomus tentans (Diptera:
Chironomidae). Chemosphere 56, 701–706.
David, J.P., Strode, C., Vontas, J., Nikou, D., Vaughan, A., Pignatelli,
Maitra, S., Dombrowski, S.M., Basu, M., Raustol, O., Waters, L.C.,
P.M., Louis, C., Hemingway, J., Ranson, H., 2005. The Anopheles
Ganguly, R., 2000. Factors on the third chromosome affect the level of
gambiae detoxification chip: a highly specific microarray to study
cyp6a2 and cyp6a8 expression in Drosophila melanogaster. Gene 248,
metabolic-based insecticide resistance in malaria vectors. Proc. Natl.
Maitra, S., Price, C., Ganguly, R., 2002. Cyp6a8 of Drosophila
Dombrowski, S.M., Krishnan, R., Witte, M., Maitra, S., Diesing, C.,
melanogaster: gene structure, and sequence and functional analysis of
the upstream DNA. Insect Biochem. Mol. Biol. 32, 859–870.
induced expression of the Cyp6a2 allele of a high producer strain of
McGraw, L.A., Gibson, G., Clark, A.G., Wolfner, M.F., 2004. Genes
CYP6A2 in the genetic background of a low producer strain. Gene
regulated by mating, sperm, or seminal proteins in mated female
Drosophila melanogaster. Curr. Biol. 14, 1509–1514.
Dunkov, B.C., Guzov, V.M., Mocelin, G., Shotkoski, F., Brun, A.,
Menzel, R., Bogaert, T., Achazi, R., 2001. A systematic gene expression
Amichot, M., Ffrench-Constant, R.H., Feyereisen, R., 1997. The
screen of Caenorhabditis elegans cytochrome P450 genes reveals CYP35
Drosophila cytochrome P450 gene Cyp6a2: structure, localization,
as strongly xenobiotic inducible. Arch. Biochem. Biophys. 395, 158–168.
heterologous expression, and induction by phenobarbital. DNA Cell
Miota, F., Siegfried, B.D., Scharf, M.E., Lydy, M.J., 2000. Atrazine
induction of cytochrome P450 in Chironomus tentans larvae. Chemo-
Feyereisen, R., 2005. Insect cytochrome P450. In: Gilbert, L.I., Iatrou, K.,
Gill, S.S. (Eds.), Comprehensive Molecular Insect Science. Elsevier,
Natsuhara, K., Shimada, K., Tanaka, T., Miyata, T., 2004. Phenobarbital
induction of permethrin detoxification and phenobarbital metabolism
Fuchs, S., Spiegelman, V.S., Belitsky, G.A., 1993. The effect of the
in susceptible and resistant strains of the beet armyworm Spodoptera
cytochrome P-450 system inducers on the development of Drosophila
exigua (Huebner). Pestic. Biochem. Physiol. 79, 33–41.
melanogaster. J. Biochem. Toxicol. 8, 83–88.
Nebert, D.W., 1991. Proposed role of drug-metabolizing enzymes:
Fuchs, S.Y., Spiegelman, V.S., Belitsky, G.A., 1994. Inducibility of
regulation of steady state levels of the ligands that effect growth,
various cytochrome P450 isozymes by phenobarbital and some other
homeostasis, differentiation, and neuroendocrine functions. Mol.
xenobiotics in Drosophila melanogaster. Biochem. Pharmacol. 47,
Nebert, D.W., 1994. Drug-metabolizing enzymes in ligand-modulated
Fulco, A.J., 1991. P450BM-3 and other inducible bacterial P450
transcription. Biochem. Pharmacol. 47, 25–37.
cytochromes: biochemistry and regulation. Annu. Rev. Pharmacol.
Oakeshott, J.G., Claudianos, C., Campbell, P.M., Newcomb, R.D.,
Russell, R.J., 2005. Biochemical genetics and genomics of insect
Gerhold, D., Lu, M., Xu, J., Austin, C., Caskey, C.T., Rushmore, T.,
esterases. In: Gilbert, L.I., Iatrou, K., Gill, S.S. (Eds.), Comprehensive
2001. Monitoring expression of genes involved in drug metabolism and
Molecular Insect Science. Elsevier, Oxford, pp. 309–381.
toxicology using DNA microarrays. Physiol. Genomics 5, 161–170.
Ottea, J.A., Plapp Jr., F.W., 1984. Glutathione S-transferase in the house
Girardot, F., Monnier, V., Tricoire, H., 2004. Genome wide analysis of
fly: biochemical and genetic changes associated with induction and
common and specific stress responses in adult Drosophila melanogaster.
insecticide resistance. Pestic. Biochem. Physiol. 22, 203–208.
Parvy, J.P., Blais, C., Bernard, F., Warren, J.T., Petryk, A., Gilbert, L.I.,
Guzov, V.M., Unnithan, G.C., Chernogolov, A.A., Feyereisen, R., 1998.
O’Connor, M.B., Dauphin-Villemant, C., 2005. A role for betaFTZ-
CYP12A1, a mitochondrial cytochrome P450 from the house fly. Arch.
F1 in regulating ecdysteroid titers during post-embryonic development
in Drosophila melanogaster. Dev. Biol. 282, 84–94.
G. Le Goff et al. / Insect Biochemistry and Molecular Biology 36 (2006) 674–682
Pedra, J.H., McIntyre, L.M., Scharf, M.E., Pittendrigh, B.R., 2004.
Tijet, N., Helvig, C., Feyereisen, R., 2001. The cytochrome P450 gene
Genome-wide transcription profile of field- and laboratory-selected
superfamily in Drosophila melanogaster: annotation, intron-exon
dichlorodiphenyltrichloroethane (DDT)-resistant Drosophila. Proc.
organization and phylogeny. Gene 262, 189–198.
Natl. Acad. Sci. USA 101, 7034–7039.
Ugazio, G., Burdino, E., Dacasto, M., Bosio, A., van’t Klooster, G.,
Plapp Jr., F.W., 1984. The genetic basis of insecticide resistance in the
Nebbia, C., 1993. Induction of hepatic drug metabolizing enzymes and
house fly: evidence that a single locus plays a major role in metabolic
interaction with carbon tetrachloride in rats after a single oral
resistance to insecticides. Pest. Biochem. Physiol. 22, 94–201.
exposure to atrazine. Toxicol. Lett. 69, 279–288.
Ranson, H., Hemingway, J., 2005. Glutathione transferases. In: Gilbert,
Vontas, J., Blass, C., Koutsos, A.C., David, J.P., Kafatos, F.C., Louis, C.,
L.I., Iatrou, K., Gill, S.S. (Eds.), Comprehensive Molecular Insect
Hemingway, J., Christophides, G.K., Ranson, H., 2005. Gene expression
Science. Elsevier, Oxford, pp. 383–402.
in insecticide resistant and susceptible Anopheles gambiae strains
Ranz, J.M., Castillo-Davis, C.I., Meiklejohn, C.D., Hartl, D.L., 2003.
constitutively or after insecticide exposure. Insect Mol. Boil. 14, 509–521.
Sex-dependent gene expression and evolution of the Drosophila
Wang, J., Kean, L., Yang, J., Allan, A.K., Davies, S.A., Herzyk, P., Dow,
transcriptome. Science 300, 1742–1745.
J.A., 2004. Function-informed transcriptome analysis of Drosophila
Reichert, K., Menzel, R., 2005. Expression profiling of five different
xenobiotics using a Caenorhabditis elegans whole genome microarray.
Wang, Q., Hasan, G., Pikielny, C.W., 1999. Preferential expression of
biotransformation enzymes in the olfactory organs of Drosophila
Rewitz, K., Styrishave, B., Andersen, O., 2003. CYP330A1 and CYP4C39
melanogaster, the antennae. J. Biol. Chem. 274, 10309–10315.
enzymes in the shore crab Carcinus maenas: sequence and expression
Waters, L.C., Zelhof, A.C., Shaw, B.J., Ch’ang, L.Y., 1992. Possible
regulation by ecdysteroids and xenobiotics. Biochem. Biophys. Res.
involvement of the long terminal repeat of transposable element 17.6 in
regulating expression of an insecticide resistance- associated P450 gene
Sabourault, C., Guzov, V.M., Koener, J.F., Claudianos, C., Plapp Jr.,
in Drosophila [published erratum appears in Proc. Natl. Acad. Sci. USA
F.W., Feyereisen, R., 2001. Overproduction of a P450 that metabolizes
1992 Dec 15;89(24):12209]. Proc. Natl. Acad. Sci. USA 89, 4855–4859.
diazinon is linked to a loss- of-function in the chromosome 2 ali-
Xu, W., Bak, S., Decker, A., Paquette, S.M., Feyereisen, R., Galbraith,
esterase (MdalphaE7) gene in resistant house flies. Insect Mol. Biol. 10,
D.W., 2001. Microarray-based analysis of gene expression in very large
gene families: the cytochrome P450 gene superfamily of Arabidopsis
Shirk, P.D., Minoo, P., Postlethwait, J.H., 1983. 20-Hydroxyecdysone
stimulates the accumulation of translatable yolk polypeptide gene
Yu, S.J., 1986. Consequences of induction of foreign compound-
transcript in adult male Drosophila melanogaster. Proc. Natl. Acad.
metabolizing enzymes in insects. In: Brattsten, L.B., Ahmad, S.
(Eds.), Molecular Aspects of Insect–Plant Interactions. Plenum, New
Snyder, M.J., 1998. Identification of a new cytochrome P450 family,
CYP45, from the lobster, Homarus americanus, and expression
Yu, S.J., 2004. Induction of detoxification enzymes by triazine herbicides
following hormone and xenobiotic exposures. Arch. Biochem.
in the fall armyworm, Spodoptera frugiperda (J.E. Smith). Pestic
Spiegelman, V.S., Fuchs, S.Y., Belitsky, G.A., 1997. The expression of
Yu, S.J., Terriere, L.C., 1974. A possible role for microsomal oxidases in
insecticide resistance-related cytochrome P450 forms is regulated by
metamorphosis and reproduction in the housefly. J. Insect Physiol. 20,
molting hormone in Drosophila melanogaster. Biochem. Biophys. Res.
Zijlstra, J.A., Vogel, E.W., Breimer, D.D., 1984. Strain-differences and
Terriere, L.C., 1983. Enzyme induction, gene amplification and insect
inducibility of microsomal oxidative enzymes in Drosophila melanoga-
resistance to insecticides. In: Georghiou, G.P., Saito, T. (Eds.), Pest
ster flies. Chem. Biol. Interact. 48, 317–338.
Resistance to Pesticides. Plenum Press, New York, pp. 265–297.
Zou, S., Meadows, S., Sharp, L., Yan, L.Y., Jan, Y.N., 2000. Genome-
Terriere, L.C., Yu, S.J., 1974. The induction of detoxifying enzymes in
wide study of aging and oxidative stress response in Drosophila
insects. J. Agric. Food. Chem. 22, 366–373.
melanogaster. Proc. Natl. Acad. Sci. USA 97, 13726–13731.
Trustees APPLICATIONS FOR 2014 SCHOLARSHIPS GUIDANCE NOTES You must read these notes carefully. A major reason for rejection is failure to provide the necessary information by the required deadline. Note: “Home institution” means the place where you are currently working “Host Institution” means the place you wish to visit The HIV Research Trust Scholarships a
Assessment of Bacterial Contamination of Raw Meats Sold in Korea, 2007 Dokyung Lee, Jaewoong Hwang, Hwanjin Yang, Soek Jang 1 , Eunhye Baek 1 , Mijin Kim 1 , Junghyun Kim 1 , Sangjin Lee 2 and Namjoo Ha ※ Department of Pharmacy, 1 Department of Life Science, 2 Division of Animal Science, Sahmyook University, Seoul 139-742, Korea 2007년 한국에서 판매된 식육의 미�

 Insect
Insect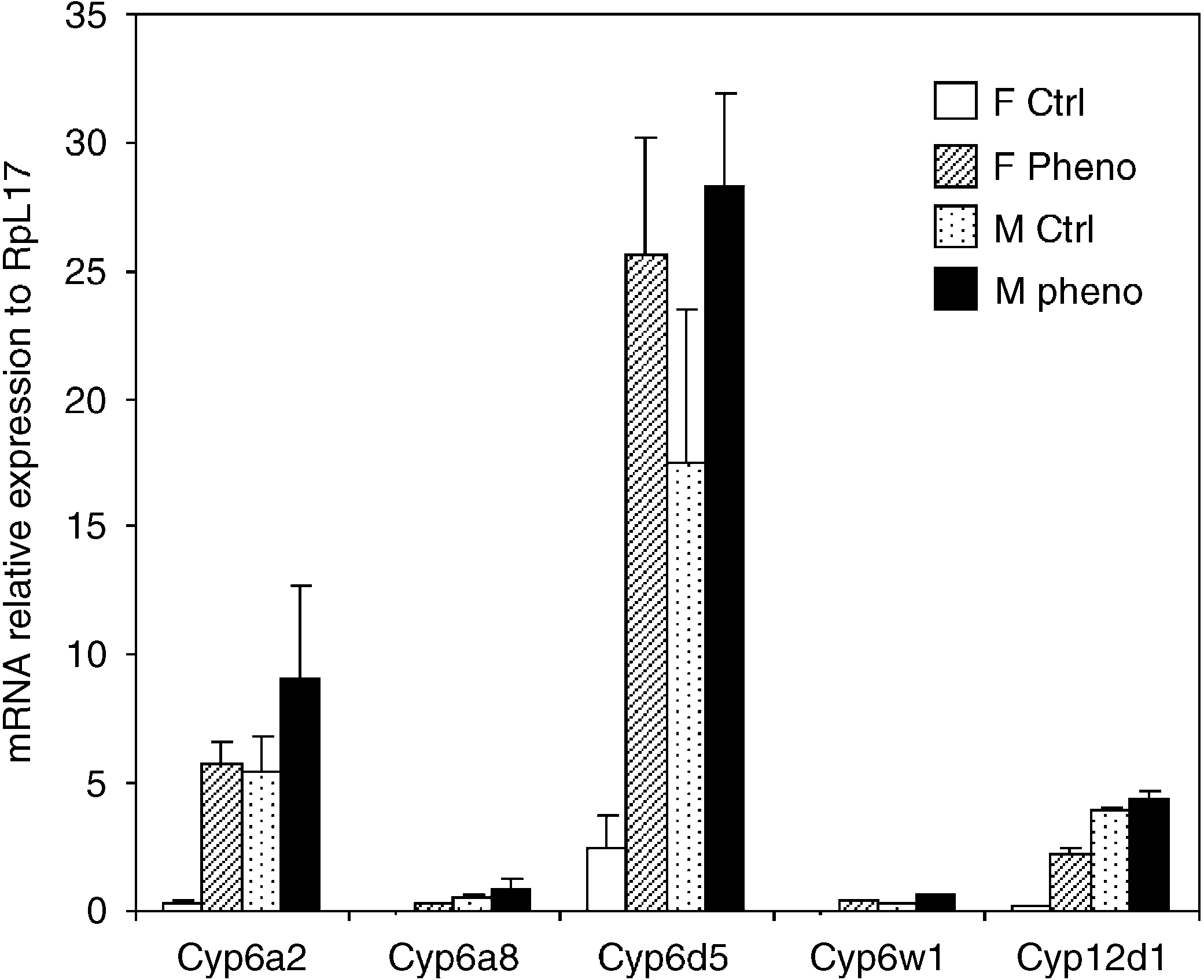
 G. Le Goff et al. / Insect Biochemistry and Molecular Biology 36 (2006) 674–682
Fig. 3. Effects of phenobarbital on yolk protein mRNA levels in adultmales of Drosophila. Validations of microarray data by qPCR. Datapresented are the ratio of YP to Rpl17.
G. Le Goff et al. / Insect Biochemistry and Molecular Biology 36 (2006) 674–682
Fig. 3. Effects of phenobarbital on yolk protein mRNA levels in adultmales of Drosophila. Validations of microarray data by qPCR. Datapresented are the ratio of YP to Rpl17.