Tadalafil appartiene alla classe degli inibitori selettivi della fosfodiesterasi di tipo 5, con un profilo farmacocinetico caratterizzato da un’emivita terminale di circa diciotto ore. Dopo somministrazione orale viene assorbito rapidamente e raggiunge concentrazioni plasmatiche massime in due ore. La biotrasformazione avviene principalmente tramite CYP3A4 con formazione di metaboliti inattivi, escreti in prevalenza con le feci. L’elevato legame alle proteine plasmatiche (>90%) assicura una distribuzione stabile. Nei confronti delle altre molecole della stessa classe, cialis compresse italia è noto per la durata prolungata dell’attività farmacologica.
Doi:10.1016/j.theriogenology.2005.05.005
www.journals.elsevierhealth.com/periodicals/the
School of Agricultural and Veterinary Sciences, Charles Sturt University, Wagga Wagga,
Genetic sperm defects are specific sperm defects, which have been shown to have a genetic mode
of transmission. Such genetic linkage, either direct or indirect, has been associated with a number ofsperm defects in different species, with this number increasing with improved diagnostic capabilities.
A number of sperm defects, which have proven or suspected genetic modes of transmission are
discussed herein, with particular emphasis on cattle. These include:
1. Acrosome defects (knobbed, ruffled and incomplete);2. Head defects (abnormal condensation, decapitated, round head, rolled head, nuclear crest);3. Midpiece abnormalities (‘‘Dag’’ defect, ‘‘corkscrew’’ defect, ‘‘pseudo-droplet’’ defect);4. Tail defects (‘‘tail stump’’ defect, primary ciliary dyskinesia).
# 2005 Elsevier Inc. All rights reserved.
Keywords: Genetic sperm defects; Male fertility; Sperm morphology
Sperm abnormalities have long been associated with male infertility and sterility in most
species studied. These abnormalities vary from morphological defects that are evident uponclinical examination, to those, which are more subtly defective. In general, sperm structurecan play a substantial role in both fertilization and pregnancy outcome The causes ofdefective sperm structure may be environmental, genetic, or a combination of both. Althoughenvironmental causes are considered to be most common, there is a growing list of spermstructural defects, which are considered to be of genetic origin.
E-mail address: [email protected].
0093-691X/$ – see front matter # 2005 Elsevier Inc. All rights reserved. doi:10.1016/j.theriogenology.2005.05.005
P.J. Chenoweth / Theriogenology 64 (2005) 457–468
Even though the heritability of bull fertility is generally considered to be low, certain
aspects of bull fertility, including sperm morphological abnormalities, are under geneticcontrol . Earlier work associated sire differences with the proportion of normalsperm, including a tendency for inbred bulls to have more morphological spermabnormalities than line-cross bulls. This was reinforced by further work with the same linesof bulls in which degree of inbreeding was associated with increased proportions ofabnormal seminiferous tubules. Much of the variation (62%) in non-return rate usingfrozen semen was associated with bulls , although this study included only AI bullspreviously screened for genital health and semen quality (which would tend to reduce theadverse effects of those variables).
A number of classification systems exist for sperm abnormalities, including the
Sperm abnormalities have been classified on the basis of their presumptive origin
. In this system, defects which occur during spermatogenesis are considered asprimary and those developing subsequent to spermiation considered as secondary. Although primary defects were originally considered to be caused by some type of directinsult to the seminiferous epithelium, they were not automatically assumed to be moredeleterious to fertility than secondary defects, which could be induced by a variety ofcauses, including iatrogenic. Later interpretations of this system included assumptions thatprimary defects are more adverse to male fertility than secondary defects , and that thelatter are more variable and less consequential than the former. These assumptions areundergoing serious review in light of recent developments in cell biology and proteomics.
The primary/secondary classification was revised as exceptions to the rule became
evident. In the revised system, sperm defects were classified as either major or minor interms of their perceived adverse effects upon male fertility. Here, major sperm defects werethose, which had proven to be associated with impaired fertility, and minor sperm defectswere those which were generally considered to be of minor consequence to male fertility. Further definition of major sperm defects (sometimes referred to as specific sperm defects)included the following criteria:
1. They are well characterized ‘‘primary’’ sperm defects,2. They occur in a substantial proportion (at least 10–15%) of the sperm population,3. They are consistent in occurrence,4. They are associated with male infertility or sterility,5. They may be heritable.
P.J. Chenoweth / Theriogenology 64 (2005) 457–468
Table 1Major and minor sperm defects in the bull
Strongly coiled or folded tail (‘‘Dag’’ defect)
A further tacit caveat is that such defects occur in the absence of a clinically or
This system was used to develop a presumptive list of major and minor sperm defects in
Considerable overlap occurs between this system (based upon known effects upon
fertility) and the primary/secondary system (based upon origin of the defect), to the extentthat lists of sperm defect categories in either system are strikingly similar. Closer scrutinyhowever suggests that a number of ‘‘minor’’ defects most probably occur prior tospermiation (e.g. subtle sperm head abnormalities and abaxial implantation). Certainly,effective use of the major/minor sperm defect system also requires critical interpretation. For example, as knowledge grows in areas of cell biology, genomics and proteomics, ourunderstanding of sperm ‘‘fertility’’ becomes more complicated and the list of ‘‘major’’sperm defects tends to increase. In addition, computer-assisted image analysis enables us toidentify subtle shape differences between sperm We now better understand that thespermatogenic epithelium responds to a wide variety of stressors in a predictable fashion,such that many sperm abnormalities previously regarded as being finite entities are nowconsidered to be different representations of the same response process. This stereotypedspermatogenic stress response is characterized, at one end of the spectrum, by the subtlediadem defect of sperm, which cascades through to gross sperm head abnormalities withincreasing severity or duration of spermatogenic stress
2.3. Compensable and uncompensable semen traits
Abnormal sperm may reduce fertility in one of two ways: (1) failure to reach the
fertilization site; or (2) inability to fertilize the ovum once they are at the fertilization site orto sustain development of the early embryo In the first case, failure of sperm to reachthe fertilization site can often be traced to problems in sperm transport. Sperm defects
P.J. Chenoweth / Theriogenology 64 (2005) 457–468
Fig. 1. The crater/diadem defect and its sequelae.
which cause either impaired sperm motility, or reduced probability of successfullytranscending the female tract, are termed compensable defects. This is because atheoretical increase in numbers of functionally competent sperm will solve, or mitigate, theproblem. Those defects which lead to failed fertilization or early pregnancy loss are termeduncompensable With these defects, an increase in sperm numbers alone willtheoretically not improve fertility.
Under natural breeding conditions, there is continuous natural selection against
inherited factors, which reduce fertility. In addition, natural barriers occur within the maleand female tract to remove faulty gametes. However, modern breeding methods,particularly those involving advanced assisted reproductive technologies (e.g. IVF, ICSI),may circumvent these principles and barriers such that genetic infertility factors may bepropagated via subfertile males In this discussion, we will focus upon genetic spermdefects where the following quotation is relevant. ‘‘Some types of defects appear in thesemen at a fairly constant rate and in a very high proportion of the sperm cells without any
P.J. Chenoweth / Theriogenology 64 (2005) 457–468
indication of environmental influence. Such defects may be presumed to be rooted in thebull’s genome and the prognosis for future improvements in semen quality would be verypoor’’
Genetic sperm defects are those that have been shown to have a genetic mode of
transmission. Such genetic linkage has been associated with a number of sperm defects indifferent species, with this number increasing with improved diagnostic capabilities. However, it should be noted that other sperm defects, more variable in nature that may beinfluenced by an interaction of environment and genetic predisposition. The elucidation ofthe genetic basis of sperm defects presents difficulties and it is no coincidence that thenumber of identified genetic sperm defects is related to the rate of adoption of AI of thespecies involved. Thus, the number of genetic sperm defects identified in cattle isconsiderably greater than in many other species. This should not, however, imply that cattleare more afflicted in this regard than are other species. Rather, they have probablyundergone more scrutiny than most. Such scrutiny should follow appropriate protocols,such as those described below, to properly establish a genetic basis for the defect inquestion. Here, the National Association of Artificial Breeders (NAAB) in USA recognizesthe importance of identifying genetic faults, as is evident by the following quotation. ‘‘Themajor responsibility of NAAB is to assure that effective programs are implemented by theAI industry. . . to control the proliferation of economically important gene defects’’. Tofirmly establish a nominate defect as having a genetic basis, the NAAB requires thefollowing questions to be answered:
1. Has evidence been provided that establishes the presence of the condition in the cattle
2. Is the mutation within germinal or somatic cells?3. What breed (or breeds) is involved?4. Is the condition lethal, and at what time in life is the condition expressed?5. What proportion of the population may be affected?6. Are similar conditions known in other species?7. Is there evidence of a selective advantage for a heterozygote?8. Are tests currently available to detect the condition?9. Are the tests definitive and what are their limitations?
Once the problem is recognized as an adverse genetic condition, it is important to
1. Its physiological basis. 2. Its mode of inheritance. 3. Its economic consequences. 4. An accurate rapid and inexpensive diagnostic test. 5. Control procedures.
Interestingly, in its deliberations to date, the NAAB has not identified any genetic sperm
defects. Rather, the current approved lists of identified genetic defects in both dairy andbeef cattle contain structural, neurological and biochemical defects only. In the following
P.J. Chenoweth / Theriogenology 64 (2005) 457–468
discussion, attempts are made to identify, in animals of veterinary interest, a number ofrelevant sperm defects, which are either known or suspected to have a genetic basis, at leaston occasion.
3. Categorization of genetic sperm defects
This sperm defect was first reported in a sterile Friesian bull and subsequently
associated with an autosomal sex-linked recessive mode of genetic transmission in thisbreed . It has been associated with infertility in bulls, boars and rams Inboars, the defect has been associated with both dominant and sex-linked recessive modes of transmission. There is some evidence that it exists as a genetic defect within theAngus breed in North America (Chenoweth unpublished) and there is a suggestion of agenetic linkage in Charolais cattle .
In the bull, the major manifestations of this abnormality are: (1) a refractile, thickened
acrosomal apex; and (2) an indented sperm apex Electron microscopy often reveals acystic region (‘‘cystic apical body’’ ) containing vesicles with inclusions, as well asabnormal fusion of acrosomal membranes . There is also often a bending back, orabrupt termination, of sperm nuclear material .
Elevated levels of knobbed acrosomes (KA) in bull semen may be caused by either
environmental or genetic factors. When environmental, they are usually transitory andassociated with other signs of spermatogenic dysfunction (i.e. increased spermabnormalities in general, including nuclear vacuoles). A genetic cause is suspected whenhigh proportions of sperm exhibit the KA defect in the absence of frequent numbers ofother sperm abnormalities, and when the defect persists at a consistently high level . InCanada, 78 of 1331 (0.53%) bulls had sperm with knobbed acrosomes (cause unknown)In contrast, in our laboratory, we found an estimated genetic prevalence of 6.74% in aknown affected Angus herd (unpublished). Sperm containing KA either lack the ability toattach to ova or have reduced capability to do so . If this were the only adverseeffect of KA, then this defect would satisfy the ‘‘compensable’’ category, wherebyincreasing sperm numbers could compensate for damaged sperm. In fact, this may helpexplain an observed difference between natural and AI fertility of a KA affected ram . In addition, the proportion of KA sperm was shown to decrease during transit in the cow’sgenital tract . Despite these findings, in vitro studies indicate that apparently normalsperm from animals affected by KA may also have compromised fertility Furtherstudies have shown that these apparently morphologically normal sperm can have plasmamembrane damage, and show premature capacitation, spontaneous acrosome reaction, andimpaired chromatin condensation . Thus the KA defect may encompass both‘‘compensable’’ and ‘‘uncompensable’’ characteristics. As bulls with apparently similarproportions of the defect may vary in infertility, this variation may be either due to suchunrecognized causes of sperm dysfunction, or to the numbers of undamaged spermreaching the fertilization site.
P.J. Chenoweth / Theriogenology 64 (2005) 457–468
Ruffled and incomplete acrosomes have been reported in subfertile bulls where they
were linked with the knobbed acrosome defect . Here, ruffled acrosomes had anirregular staining pattern leading to a wrinkled, or ruffled, appearance. Incompleteacrosomes had an irregular margin; giving the appearance that part of the acrosome wasmissing or incomplete. A genetic basis was suggested by the occurrence of the three defects(knobbed, ruffled and incomplete acrosomes) in four sons of a subfertile Holstein sire. Some similarity has also been drawn with acrosome abnormalities described in‘‘genetically-determined quasi-sterile’’ male mice
‘‘Differences in spermatozoan DNA exist, not only among individuals of the same
species but also among sperm in the same ejaculate’’ a reference related to Fuelgen-DNA staining patterns of sperm. Abnormal sperm DNA condensation (‘‘clumping’’) isdifficult to identify using routine sperm morphology techniques. However, the use of flowcytometry, in concert with DNA-specific fluorochromes, was effective in detecting thedegree of heterogeneity of sperm nuclear chromatin structure; this was associated withdisturbances of spermatogenesis, sperm abnormalities and infertility in a number of speciesIt is included in this discussion as it may provide an example of genetic-environmental interactions on sperm structure. A Canadian Simmental bull identified withthis condition showed variable levels of infertility as well as DNA ‘‘clumping’’, with thelatter appearing to increase over the summer months .
3.2.2. Decapitated (disintegrated) sperm defect
Separation of the sperm head and tail can be caused by a number of adverse factors
affecting either spermiogenesis or sperm maturation However, a specific, sterilizingform has been reported in several cattle breeds (Guernsey, Hereford, Swedish Red &White) where it has been associated with sterility. Most (80–100%) of sperm are affected,with the separated tail usually remaining motile. In addition, the proximal end of theseparated midpiece is often curled around a cytoplasmic droplet, giving the appearanceperhaps of a micro-cephalic sperm head. The separation has been associated with defectivedevelopment of the sperm head, implantation grove and basal plate, and becomes evidentwhen sperm are traversing the epididymis Evidence for the hereditary nature ofthis defect in bulls (most probably via a sex-limited recessive gene with male and femalecarriers) has come from several reports A similar defect was reported in a group(n = 8) of Hereford bulls in which testicular hypoplasia (n = 5) was also diagnosed.
The following characteristics have been proposed as being indicative of this specific
1. Sperm heads and tails are separated in 80–100% of ejaculated sperm,2. A high percentage of the loose tails show active movement,3. A proximal bending or curling of the middle-piece around the cytoplasmic droplet is
P.J. Chenoweth / Theriogenology 64 (2005) 457–468
This defect has been reported in four infertile men (two of whom were brothers), where
it affected 100% of ejaculated sperm Many of the sperm heads contained vacuole-likestructures, while none had an acrosome attached.
3.2.4. Rolled-head, nuclear crest, giant head syndrome
A combination of abnormalities (rolled-head, nuclear crest and giant heads) have been
observed to occur in combination in some bulls. Where such abnormalities occur insubstantial numbers, there have been suggestions of hereditary linkages . Theultrastructure of sperm with rolled heads and nuclear crests has been described It issuggested that rolled sperm represent a deviated form of the giant sperm headabnormality and that they are often diploid (and occasionally triploid or eventetraploid) Bulls with this defect generally exhibit a consistent spermiogramover time Both the nuclear crest and rolled head defects were also reported to becaused by ethylene-dibromide spermatoxicity Although effects of this sperm defectupon fertility are unclear it is logical to assume that abnormal chromosome numbersin sperm would compromise fertility, as encountered in males with Kleinfelter’ssyndrome
Named after the Jersey bull in which the defect was first identified, this defect is
represented by strong folding, coiling and fracture of the distal part of the sperm midpiece(with or without a retained distal cytoplasmic droplet). A consistent similar spermiogramwas observed in a full brother to ‘‘Dag’’, suggesting a hereditary basis. However, anenvironmental link with dietary zinc has also been suggested The defect may reflectdisturbance in the testis or epididymis and may be present (<4%) in normal semen. Levelsabove 50% can have serious fertility implications A similar defect has been reportedin a subfertile boar , although a genetic basis was not established.
This was first reported in five Friesian bulls in Denmark where the defect was
encountered in 7–26% of ejaculated sperm. As the affected bulls were all related (two werehalf-brothers), a hereditary basis was suggested. The semen samples from the affected bullswere usually of normal concentration, although initial sperm motility was poor. Thepercentage of affected sperm tended to increase with bull age.
The defect was characterized by a local thickening on the midpiece. Although they
could resemble cytoplasmic droplets under normal microscopy, they are prone to occur inregions where droplets are seldom encountered (e.g. the middle of the midpiece). They arealso more likely to be irregular in shape and more visually dense than droplets. Ultra-microscopically, they comprised accumulations of granules surrounded by mitochondria.
A ‘‘mictotubular mass defect’’ of spermatozoa was reported in the semen of seven
Standardbred stallions, in which a genetic link was suggested . Here, irregular massesin the proximal region of the midpiece contained torturous arrays of small abnormal
P.J. Chenoweth / Theriogenology 64 (2005) 457–468
microtubules. Similar structural defects (termed ‘‘knobs’’) were observed in a sterilestallion , where they represented mitochondrial accumulations.
Similar irregular midpiece formations in bull semen have been associated with Bovine
Ephemeral Fever and also with gossypol spermatoxicity In the latter case, thesemasses were also caused by abnormal accumulations and clumping of mitochondria.
3.3.3. ‘‘Corkscrew midpiece’’ defect
The ‘‘corkscrew sperm defect’’ was first described in ejaculated sperm of two sterile
bulls where it was observed as an irregular distribution (‘‘lumps’’) of mitochondria,resembling a corkscrew under light microscopy. A genetic cause was suggested as four ofthe first five detected bulls were related. However, similar defects have been reported inbulls following Bovine Ephemeral Fever , in a sterile stallion and in bullsundergoing gossypol spermatoxicity Further doubt concerning the genetic basis ofthis defect came from the original source, who associated an observed temporal increase ofthis defect in Danish bulls with concurrent increased levels of radioactive fallout Here, an environmental-genetic interaction may have been evident as the Red Danish breedappeared to be particularly susceptible to the spermatoxic effect in question.
Simple coiled tails (a term now understood to include the distal midpiece reflex) are
among the most common sperm defects. Often, an increased prevalence of such defects inthe ejaculate is associated with a one or more of a variety of non-genetic origin etiologies.
This defect was first reported in bulls in 1925 It has also been encountered in the
mouse, rabbit, dog, stallion and man Three sterile Canadian bulls (Ayrshire,Shorthorn, Holstein) showed virtual absence of the sperm midpiece and tail in the majorityof ejaculated sperm, where they were represented by a small ‘‘stump’’ or ‘‘stub’’ Inaddition, sperm concentration was very low and sperm motility was virtually absent. Monitoring of sperm morphology in these bulls indicated that the percentage of sperm withthe defect increased with the age of the bulls. It has been suggested that a prevalence ofgreater than 25% of this defect in the ejaculate is suggestive of a genetic fertility problemOther reports have linked this defect with sterility in bulls and suggestion hasbeen made of an inherited mode of transmission . It has also been linkedwith infertility in boars and reported in man as well as mice. Care should be takento differentiate this condition from the ‘‘accessory tail defect’’ in bulls, which probablyshares a common etiology with abaxial sperm midpieces, as well as little impact uponfertility
3.4.3. Primary ciliary dyskinesia (previously termed immotile cilia syndrome)
This is represented by a diverse group of disorders characterized by a structural and
generalized abnormality of ciliated cells. In man, a systemic axonemal alteration isassociated with Kartagener’s syndrome in which the males are infertile and possess
P.J. Chenoweth / Theriogenology 64 (2005) 457–468
immotile spermatozoa . Here, in common with other ciliated cells in the body (such asrespiratory epithelial cells) affected sperm (and respiratory tract cilia) have perturbedaxonmenal structures, e.g. part or complete absence of dynein arms, microtubuledisorganization, or absent radial spokes . This suggests a genetic link between thesestructures or that they are coded by the same gene. Similar sperm aberrations exist in theanimal world, although the link with respiratory diseases has not been adequately pursued. It should be noted that the axonemal complex contains over 200 proteins, defects in any ofwhich could be related to problems with genetic coding. Similar alterations to sperm tailaxonemal complexes have been identified in genetically similar mice, which were sterile
[1] Williams WW, Savage A. Observations on the seminal micropathology of bulls. Cornell Vet 1925;15:
[2] Saacke RG, White JM. Semen quality tests and their relationship to fertility. In: Proceeding of the 4th NAAB
Tech Conference on AI and Reprod. 1972. p. 22–7.
[3] Wiltbank JN, Parrish NR. Pregnancy rates in cows and heifers bred to bulls selected for semen quality.
[4] Barth AD. Influence of abnormal sperm morphology on conception rates in cattle. Proc Soc Theriogenol
[5] Chandler JE, Adkinson RW. Genetic and non-genetic contributions to variation in spermatozoal morphology
of mature Holstein bulls. In: Proceeding of the NAAB 13th Technical Conference on AI and Reprod. 1990. p. 57–62.
[6] Zamboni L. Sperm structure and its relevance to infertility. Arch Pathol Lab Med 1992;116:325–44. [7] Saacke RG. What is a BSE–SFT standards: the relative importance of sperm morphology: an opinion. Proc
[8] Salisbury GW, Baker FN. Nuclear morphology of spermatozoa from inbred and linecross Hereford bulls.
[9] Carroll EEJ, Ball L. Testicular changes as affected by mating systems in beef cattle. Am J Vet Res
[10] Milovanov VK. Artificial insemination of farm animals (Russ) 5 ed, 1940. Moscow. [11] Blom E. Interpretation of spermatic cytology of bulls. Fert Steril 1950;1:223–38. [12] Barth AD, Oko RJ. Abnormal morphology of bovine spermatozoa. Ames, IA: Iowa State Press (Blackwell),
[13] Blom E. The ultrastructure of some characteristic sperm defects and a proposal for a new classification of the
bull spermiogram. In: Atti del VII Simposio Int di Zootechnia Milan. 1972. p. 125–39.
[14] Ostermeier GC, Sartor-Bergfelt R, Susko-Parrish JL, Parrish JJ. Bull infertility and sperm nuclear shape. Ag
[15] Saacke RG. What is abnormal? And is abnormal dependent upon the animal.In: Proceeding of the NAAB
13th Technical Conference on AI and Reprod. 1990. p. 67–73.
[16] Saacke RG, Amann RP, Marshall CE. Acrosomal cap abnormalities of sperm from subfertile bulls. J Anim
[17] Teunissen GHP. Een afwijkning van het acrosom (kopcap) bij de spermatozoiden van een stier. Tijfdschr
[18] Donald HP, Hancock JL. Evidence of a gene-controlled sterility in bulls. J Agric Sci (Camb) 1953;
[19] Barth AD. The knobbed acrosome defect in beef bulls. Can Vet J 1986;27:379–84. [20] Soderquist L. Reduced fertility after artificial insemination in a ram with a high incidence of knobbed
acrosomes. Vet Rec 1998;143:227–8.
P.J. Chenoweth / Theriogenology 64 (2005) 457–468
[21] Wohlfarth E. Beitrag zum akrosom-defekt im eberspermia. Zuchthyg FortpflStor Besam Haustiere
[22] Bishop MWH. Genetically determined abnormalities of the reproductive system. J Reprod Fert Suppl
[23] Thundathil J, Meyer R, Palasz AT, Barth AD, Mapletoft RD. Effect of knobbed acrosome defect in bovine
semen on IVF and embryo production. Theriogenology 2000;54:921–34.
[24] Bane A, Nicander L. Electron and light microscopical studies on spermateliosis in a boar with acrosomal
abnormalities. J Reprod Fert 1966;11:133–8.
[25] Cran DG, Dott HM. The ultrastructure of knobbed bull spermatozoa. J Reprod Fert 1976;47:407–8. [26] Buttle HRL, Hancock JL. Sterile boars with ‘‘knobbed’’ spermatozoa. J Agric Sci (Camb) 1963;65:
[27] Mitchell JR, Senger PL, Rosenberger JL. Distribution and retention of spermatozoa with acrosomal and
nuclear abnormalities in the cow genital tract. J Anim Sci 1985;61:956–67.
[28] Thunderthil J, Palasz AT, Barth AD, Mapletoft RJ. Plasma membranes and acrosomal integrity in bovine
spermatozoa with the knobbed acrosome effect. Theriogenology 2002;58:97–102.
[29] Rajasekarasetty MR. Studies on a new type of genetically determined quasi-sterility in the house mouse. Fert
[30] Salisbury GW, Hart RG, Lodge JR. The spermatozoan genome and fertility. Am J Ob Gyn 1977;128:342–50. [31] Ballachey RE, Hohenboken WD, Evanson DP. Heterogeneity of sperm nuclear chromatin structure and its
relationship to bull fertility. Biol Reprod 1987;36:915–25.
[32] Settergren I, Nicander L. Ultrastructure of disintegrated bull sperm. In: Proceeding of the 6th International
Congress Animal Reprod and AI, vol. 1; 1968.p. 191.
[33] Blom E, Birch-Andersen A. Ultrastructure of the decapitated sperm defect in Guernsey bulls. J Reprod Fert
[34] Jones WA. Abnormal morphology of the spermatozoa in Guernsey bulls. Br Vet J 1962;118:257. [35] Van Rensburg SWJ, Van Resnburg SJ, de Vos WH. The significance of the cytoplasmic droplet in the
disintegration of semen in Guernsey bulls. Onderstepoort J vet Res 1966;33:169.
[36] Milk Marketing Board (UK). Decapitated sperm. Rep Breed and Prod Org Milk Market Brd 1967/
[37] Williams G. An abnormality of the spermatozoa of some Hereford bulls. Vet Rec 1965;77:1204–6. [38] Blom E. A decapitated sperm defect in two sterile Hereford bulls. Nord Vet-Med 1977;29:119–23. [39] Kullander S, Rausing A. On round-headed human spermatozoa. Int J Fert 1975;20:33–40. [40] Cran DG, Dott HM, Wilmington HW. The structure and formation of rolled and crested bull spermatozoa.
[41] Blom E. Rolled head and nuclear crest sperm defects in a rare case of presumed diploidy in the bull. Nord
[42] Gledhill BL. Cytophotometry of presumed diploid bull spermatozoa. Nord Vet Med 1965;17:328–35. [43] Eljack AH, Hrudka F. Patterns and dynamics of teratospermia induced in rams by parenteral treatment with
ethylene dibromide. J Ultrastruct Res 1979;7:124–34.
[44] Lanfranco F, Kamischke A, ZitzmannM. Nieschlag E. Kleinfelter’s syndrome. Lancet 2004;364:273–83. [45] Blom E, Wolstrup C. Zinc as a possible causal factor in the sterilizing sperm tail defect, the ‘‘Dag
defect’’, in Jersey bulls. In: Proceeding of the VIIIth International Congress Anim Reprod and AI, vol. IV;1976.p. 690–3.
[46] van Duijn C. Ultrastructural mid-piece defects in spermatozoa from the subfertile Great Yorkshire boar Ard.
In: Proceeding of the 7th International Congress Anim Reprod and AI, vol. 1. 1972. p. 469–73.
[47] Blom E. A new sperm defect: pseudo-droplets, in the middle piece of bull sperm. Nord Vet-Med
[48] Heath E, Aire T, Fujiwara K. Microtubular mass defect of spermatozoa in stallions. Am J Vet Res
[49] Chenoweth PJ, Pascoe RR, McDougall HL, McCosker PJ. An abnormality of the spermatozoa of a stallion
(Equus caballus). Br Vet J 1970;126:476–88.
[50] Burgess GW, Chenoweth PJ. Mid-piece abnormalities in bovine semen following experimental and natural
cases of bovine ephemeral fever. Br Vet J 1975;131:536–44.
P.J. Chenoweth / Theriogenology 64 (2005) 457–468
[51] Chenoweth PJ, Chase Jr CC, Risco CA, Larsen RE. Characterization of gossypol-induced sperm abnorm-
alities in bulls. Theriogenology 2000;53:1193–203.
[52] Blom E. A rare sperm abnormality: corkscrew-sperm associated with sterility in bulls. Nature 1959;
[53] Blom E. The corkscrew sperm defect in Danish bulls: a possible indicator of nuclear fallout? Nord Vet-Med
[54] Courbrough RI, Barker CAV. Spermatozoa: an unusual middle piece abnormality associated with sterility in
bulls. In: Proceeding of the 5th International Congress Anim Reprod and AI; 1964.p. 219–29.
[55] Blom E. A sterilizing ‘‘tail stump sperm defect’’ in a Holstein Friesian bull. Nord Vet-Med 1976;28:295–8. [56] Williams G. Tail stump defect affecting the spermatozoa of two Charolais bulls. Vet Rec 1987;121:248–50. [57] Peet R, Mullins K. Sterility in a poll Hereford bull associated with the tail stump sperm defect. Aust Vet J
[58] Vierula M, Alanko M, Andersson M, Vanhaperttula T. Tail stump defect in Ayrshire bulls: morphogenesis of
the defect. Andrologia 1987;9:207–16.
[59] Foote R, Hough S, Johnson L, Kaproth M. Electron microscopy and pedigree study in an Ayrshire bull with
tail-stump sperm defects. Vet Rec 1992;130:578–9.
[60] Andersson M, Makipaa R. Length of the sperm tail in fertile boars of different breeds and in sterile Yorkshire
boars affected with the hereditary ‘‘short tail’’ sperm defect. In: Proceeding of the 14th InternationalCongress Anim Reprod, vol. 2; 2000.p. 31–96.
[61] Barth A. Abaxial tail attachment of bovine spermatozoa and its effect on fertility. Can Vet J 1989;30:656–62. [62] Leestma JE, Sepsenwol S. Sperm tail axoneme alterations in the Wobbler mouse. J Reprod Fert
[63] Bacetti B, Burrini AG, Palluni V, Maver A, Reneiri T. 9 + 0 immotile spermatozoa in an infertile male.
GUIA UREA NITROGENO UREICO UREA/NITROGENO UREICO 1. DEFINICION La urea se sintetiza en el hígado a partir del amoniaco y se excreta por el riñón, que es el principal producto terminal del metabolismo de la proteína. El nitrógeno ureico en sangre (BUN) refleja el ingreso de proteínas y la capacidad excretora del riñón. 2. OBJETIVO Medir el nivel de urea en la s


 www.journals.elsevierhealth.com/periodicals/the
School of Agricultural and Veterinary Sciences, Charles Sturt University, Wagga Wagga,
Genetic sperm defects are specific sperm defects, which have been shown to have a genetic mode
of transmission. Such genetic linkage, either direct or indirect, has been associated with a number ofsperm defects in different species, with this number increasing with improved diagnostic capabilities.
www.journals.elsevierhealth.com/periodicals/the
School of Agricultural and Veterinary Sciences, Charles Sturt University, Wagga Wagga,
Genetic sperm defects are specific sperm defects, which have been shown to have a genetic mode
of transmission. Such genetic linkage, either direct or indirect, has been associated with a number ofsperm defects in different species, with this number increasing with improved diagnostic capabilities.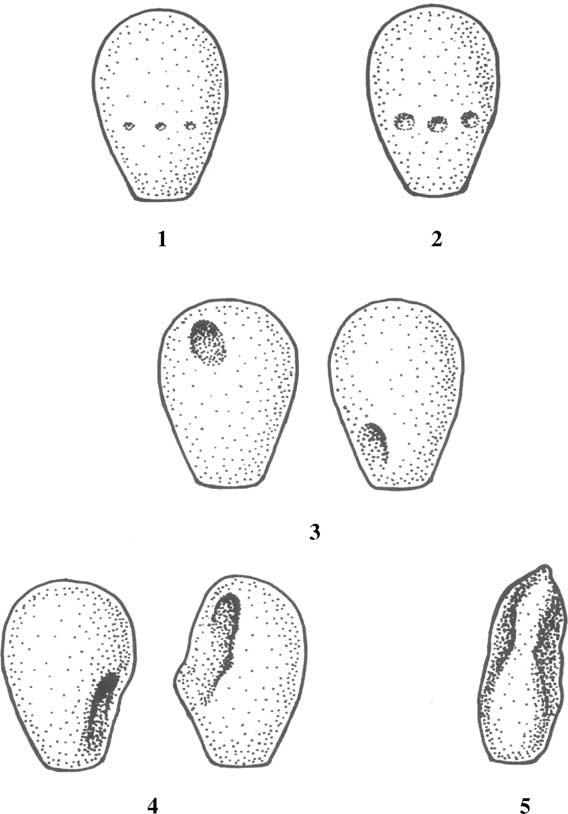 P.J. Chenoweth / Theriogenology 64 (2005) 457–468
Fig. 1. The crater/diadem defect and its sequelae.
P.J. Chenoweth / Theriogenology 64 (2005) 457–468
Fig. 1. The crater/diadem defect and its sequelae.