Tadalafil appartiene alla classe degli inibitori selettivi della fosfodiesterasi di tipo 5, con un profilo farmacocinetico caratterizzato da un’emivita terminale di circa diciotto ore. Dopo somministrazione orale viene assorbito rapidamente e raggiunge concentrazioni plasmatiche massime in due ore. La biotrasformazione avviene principalmente tramite CYP3A4 con formazione di metaboliti inattivi, escreti in prevalenza con le feci. L’elevato legame alle proteine plasmatiche (>90%) assicura una distribuzione stabile. Nei confronti delle altre molecole della stessa classe, cialis compresse italia è noto per la durata prolungata dell’attività farmacologica.
Ns140005476p
The Journal of Neuroscience, July 15, 2000, 20(14):5476–5482
Increased Excitability of Aged Rabbit CA1 Neurons after Trace Eyeblink Conditioning James R. Moyer Jr, John M. Power, Lucien T. Thompson, and John F. Disterhoft Department of Cell and Molecular Biology and the Institute for Neurosciences, Northwestern University Medical School,Chicago, Illinois 60611-3008
Cellular properties of CA1 neurons were studied in hippocampal
tential, membrane time constant, neuron input resistance, or
slices 24 hr after acquisition of trace eyeblink conditioning in
action potential characteristics. Furthermore, comparisons be-
young adult and aging rabbits. Aging rabbits required signifi-
tween CA1 neurons from trace-conditioned aging and trace-
cantly more trials than young rabbits to reach a behavioral crite-
conditioned young adult rabbits revealed no statistically signifi-
rion of 60% conditioned responses in an 80 trial session. Intra-
cant differences in postburst AHPs or accommodation, indicating
cellular recordings revealed that CA1 neurons from aging control
that similar levels of postsynaptic excitability were attained dur-
rabbits had significantly larger, longer lasting postburst afterhy-
ing successful acquisition of trace eyeblink conditioning, regard-
perpolarizations (AHPs) and greater spike frequency adaptation
less of rabbit age. These data represent the first in vitro demon-
(accommodation) relative to those from young adult control rab-
stration of learning-related excitability changes in aging rabbit
bits. After learning, both young and aging CA1 neurons exhibited
CA1 neurons and provide additional evidence for involvement of
increased postsynaptic excitability compared with their respec-
changes in postsynaptic excitability of CA1 neurons in both
tive age-matched control rabbits (naive and rabbits that failed to
learn). Thus, after learning, CA1 neurons from both age groupshad reduced postburst AHPs and reduced accommodation. No
Key words: aging; afterhyperpolarization; spike frequency ad-
learning-related differences were seen in resting membrane po-
aptation; associative learning; hippocampus; in vitro; intracellular
Aged animals, including humans, are impaired in a variety of
Landfield, 1990). In addition, calcium-dependent synaptic plasticity
learning tasks (Zyzak et al., 1995; Thompson et al., 1996). We
is altered in aging hippocampal neurons (Norris et al., 1996, 1998;
adopted the rabbit eyeblink preparation as a model system in which
Shankar et al., 1998), suggesting that one of the consequences of
to study neurobiological correlates of aging and associative learning
brain aging in mammals may be an impaired ability to modulate
(Disterhoft et al., 1977; Akase et al., 1989; Moyer et al., 1990;
intracellular calcium (Landfield, 1987; Disterhoft et al., 1994a;
Thompson et al., 1992, 1996a; McEchron and Disterhoft, 1999).
Khachaturian, 1994; Thibault and Landfield, 1996).
Aging rabbits require significantly more trials to learn trace eye-
Blockade of excess calcium influx has been shown to ameliorate
blink conditioning than young adult rabbits (Graves and Solomon,
age-related learning deficits. For example, the dihydropyridine
1985; Thompson et al., 1996a). Unlike standard delay conditioning
calcium channel antagonist nimodipine facilitates acquisition of
(Akase et al., 1989), trace eyeblink conditioning depends not only
trace eyeblink conditioning in aging rabbits (Deyo et al., 1989;
on brainstem–cerebellar circuitry but also on an intact hippocam-
Straube et al., 1990; Kowalska and Disterhoft, 1994). Intravenous
pus for successful acquisition (Moyer et al., 1990; Kim et al., 1995).
administration of the same dose of nimodipine that facilitates
Trace eyeblink conditioning deficits exhibited by aging rabbits
learning also increases spontaneous firing rates of aging rabbit CA1
parallel the deficits observed in hippocampectomized adult rabbits;
neurons in vivo (Thompson et al., 1990). Subsequent in vitro studies
both groups are profoundly impaired and show inappropriate tim-
showed that postsynaptic excitability of aging CA1 neurons can be
ing of the few conditioned responses (CRs) elicited during training
restored to levels more closely resembling young adult neurons by
(Moyer et al., 1990; Thompson et al., 1996a). These data suggest
bath application of nanomolar concentrations of nimodipine
that at least part of the deficit exhibited by aging rabbits involves
(Moyer et al., 1992; Moyer and Disterhoft, 1994). Together, these
data suggest that reducing calcium influx in aging CA1 neurons not
Hippocampal slices are a valuable tool for studying various
only alters their electrophysiological properties but also facilitates
aspects of cellular neurophysiology. Using intracellular recordings,
the ability of aged animals to learn.
we previously demonstrated that aging rabbit CA1 neurons had
Previous studies have demonstrated that ion channels can be
both larger postburst afterhyperpolarizations (AHPs) (Moyer et
modulated by associative learning (Alkon, 1984; Disterhoft et al.,
al., 1992) and prolonged calcium action potentials (APs) (Moyer
1986; de Jonge et al., 1990; Woody et al., 1991; Moyer et al., 1996;
and Disterhoft, 1994) than young adult neurons. These differences
Thompson et al., 1996b; Saar et al., 1998). For example, learning-
in calcium-mediated processes are similar to those observed in
specific reductions of the calcium-dependent slow AHP have been
aging rat CA1 neurons (Landfield and Pitler, 1984; Pitler and
observed in CA1 and CA3 neurons after acquisition of hippocam-
pally dependent trace eyeblink conditioning (Moyer et al., 1996;
Received March 6, 2000; revised April 18, 2000; accepted April 25, 2000.
Thompson et al., 1996b). Furthermore, the reduced AHPs ob-
This work was supported by National Institutes of Health Grants RO1 MH47340,
served after learning are transient, decaying back to baseline within
RO1 AG08796, and RO1 DA07633 to J.F.D. We thank F. Cutting and J. Hauser for
5–7 d, a time period appropriate for memory consolidation (Moyer
Correspondence should be addressed to Dr. James R. Moyer, Jr., Department of
et al., 1996; Thompson et al., 1996b). Similarly, reduced AHPs
Psychology, Yale University, P.O. Box 208205, New Haven, CT 06520-8205. E-mail:
were observed in layer II pyramidal neurons of rat piriform cortex
after acquisition of an odor discrimination task (Saar et al., 1998).
Dr. Power’s present address: Department of Neuroscience, Australian National
To date, no studies have used intracellular recordings in vitro to
University, John Curtin School of Medical Research, Canberra, Australia 2601.
Dr. Thompson’s present address: School of Human Development, GR 4.1, Univer-
evaluate learning-related changes in postsynaptic excitability of
sity of Texas at Dallas, Richardson, TX 75083.
CA1 neurons in aging animals. To evaluate whether learning-
Copyright 2000 Society for Neuroscience 0270-6474/00/205476-07$15.00/0
related changes also occur in aging animals, intracellular current-
Moyer et al. • Increased Excitability of Aged CA1 after Learning
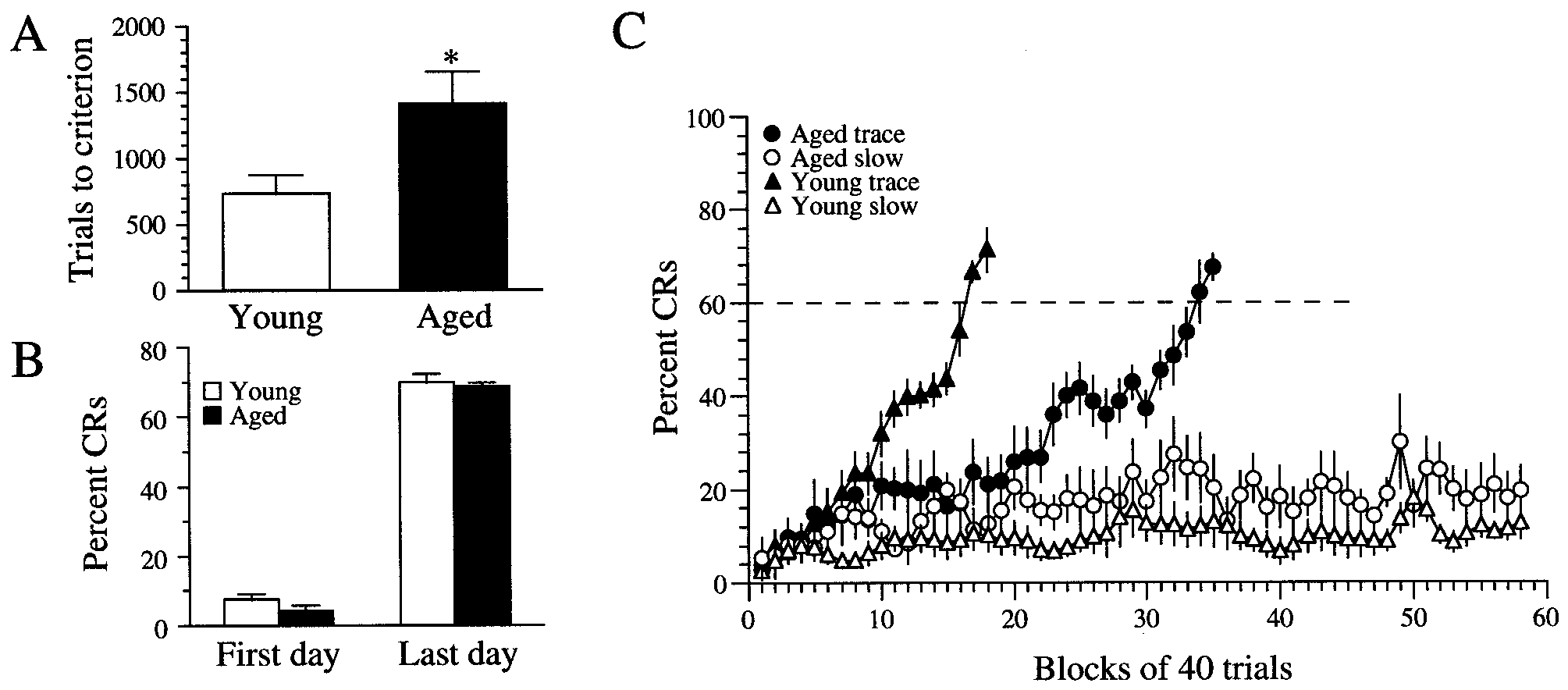
J. Neurosci., July 15, 2000, 20(14):5476–5482 5477 Figure 1. Aging rabbits are significantly impaired in their ability to learn hippocampally dependent trace eyeblink conditioning. A, A plot of trial to
criterion among rabbits that learned illustrates that aging rabbits required significantly more trials than young adult rabbits ( p Ͻ 0.05). B, A plot of percent
CRs shows that, among rabbits that learned, young and aging rabbits were not significantly different from each other on either the first or last day of training. C, Learning curves of rabbits that learned ( filled symbols) and rabbits that did not learn (open symbols). The dashed line represents the behavioral criterion
of 60% CRs. The aging rabbits that eventually learned ( filled circles; n ϭ 7) had an average learning curve that was shifted far to the right of young adult
rabbits that learned ( filled triangles; n ϭ 6). Notice the relatively poor performance of the slow-learning young adult (open triangles; n ϭ 4) and aging (opencircles; n ϭ 5) rabbits.
clamp recordings were made from CA1 pyramidal neurons in slices
For recording, slices were individually transferred to a submersion cham-
taken from aging rabbits after acquisition of trace eyeblink condi-
ber (Medical Systems, Greenvale, NY) and continuously perfused with
tioning. These data were compared with data obtained from aging
Electrophysiological recordings and data analysis. Intracellular recordings
control rabbits that did not learn, from aging naive rabbits, and
were made from 97 CA1 pyramidal neurons (50 young, 47 aging) using an
similar data from young adult rabbits.
Axoclamp 2A amplifier (Axon Instruments, Foster City, CA) and thin-
Parts of this paper have been published previously in abstract
walled microelectrodes filled with 3 M KCl (20 –50 M⍀) as described
previously (Moyer et al., 1996). All CA1 pyramidal neurons included in
this study exhibited little spontaneous activity at rest, had action potential
amplitudes Ͼ80 mV from threshold, had action potential durations Ͼ1.2
MATERIALS AND METHODS
msec from rise threshold to recrossing of the resting potential, had input
Behavioral training. New Zealand albino rabbits (Oryctolagus cuniculus)
resistances Ն25 M⍀, and had stable resting membrane potentials more
were purchased from Hazelton Rabbitry (Denver, PA) and maintained in
negative than Ϫ60 mV. Cells were studied at membrane potentials near
accordance with guidelines established by the United States Department of
Ϫ65 mV (using Յ0.2 nA constant current injection, if necessary) to
Agriculture and approved and managed by the Animal Care Committee of
minimize the influence of voltage-dependent changes on membrane con-
Northwestern University. Rabbits received 500 msec trace eyeblink con-
ductances. The protocols used to study the properties of CA1 neurons and
ditioning as described previously (Moyer et al., 1990; Thompson et al.,
the analyses of all intracellular data were identical with previously pub-
1996a). Briefly, rabbits were fitted with restraining head bolts and trained
lished methods (Moyer et al., 1996). Briefly, current–voltage relationships
in pairs in individual sound-attenuated chambers for daily 80 trial sessions
were constructed using 400 msec current injections (range, Ϫ1.0 to ϩ0.2
(mean intertrial interval, 45 sec). The CS was a 100 msec, 85 dB, 6 kHz
nA). Postburst AHPs were evaluated using a 100 msec depolarizing cur-
tone presented via stereo headphones. The unconditioned stimulus (US)
rent injection sufficient to elicit a burst of four action potentials. Spike
was a 150 msec, 3.5 psi corneal air puff sufficient to elicit reliable extension
frequency adaptation (referred to as accommodation in the present study)
of the nictitating membrane (NM) (or third eyelid) as the unconditioned
was evaluated by injecting the same amount of depolarizing current used
response. Because aging rabbits typically fail to acquire this trace eyeblink
to study the AHP but for an 800 msec duration. The number of action
conditioning task to our usual criterion of 80% CRs in a training session
potentials were counted and recorded. To evaluate the distribution of
(Thompson et al., 1996a), a behavioral criterion of 60% CRs in a training
changes within a population, individual cells were classified as having been
session was used for both age groups (all references to learning in the text
“changed by conditioning” if its data fell beyond 2 SDs from the mean of
refer to acquisition of 60% CRs unless explicitly indicated otherwise). An
the population of naive neurons studied in the particular age group (for
NM extension was counted as a CR if it occurred after CS onset but before
US onset. Slow-learning rabbits (Ͻ30% CRs after 15 sessions) served as an
All reported values are the mean Ϯ SEM. Statistical analyses were done
additional control population (Disterhoft et al., 1988a; Moyer et al., 1996;
using unpaired t tests or ANOVA with the significance level set at 0.05.
Thompson et al., 1996b). Only one young adult rabbit was slow-learning
Post hoc comparisons were made using Fisher’s PLSD only if a significant
(compared with five aging slow-learning rabbits), so three additional slow-
learning young adult rabbits were taken from a simultaneous cohort of
behavioral studies and included in the present study. Learning curves for
young and aged rabbits were normalized to the mean number of trials
required to reach criterion for that group using linear interpolation algo-
Aging rabbits are impaired in acquisition of trace
rithms (IgorPro; WaveMetrics, Lake Oswego, OR) (Thompson et al.,
1996). Behavioral experiments were controlled by an IBM-PC clone com-
eyeblink conditioning
puter using custom hardware and software as described previously (Akase
Aging rabbits required 1405 Ϯ 246 trials to reach the behavioral
et al., 1994; Thompson et al., 1994).
criterion of 60% CRs within an 80 trial trace conditioning session
Slice preparation. Twenty-four hours after the last training session,
rabbits were deeply anesthetized with halothane, and 400 m hippocampal
compared with 733 Ϯ 138 trials for young adult rabbits (t11 ϭ 2.276;
slices were cut on a vibratome as described previously (Moyer et al., 1996;
p Ͻ 0.05) (Fig. 1A). Comparisons between trace-conditioned
Thompson et al., 1996a). For this study, 17 young adult rabbits (mean age,
young and aging rabbits revealed no statistically significant differ-
2.2 Ϯ 0.1 months) and 19 aging rabbits (mean age, 42.3 Ϯ 1.3 months) were
ences in percent CRs on the first (young, 7.5 Ϯ 1.9; aging, 4.1 Ϯ 1.9;
used. Hippocampal slices were maintained in a holding chamber filled with
11 ϭ Ϫ1.27; p ϭ 0.23) or the last ( young, 69.8 Ϯ 2.6; aging, 68.6 Ϯ
2PO4, 2.4 CaCl2, 26 NaHC O3, and 10 D-glucose, gassed with 95%
11 ϭ Ϫ0.439; p ϭ 0.67) day of training. Learning curves
O2–5% CO2 at pH 7.4) at room temperature (ϳ23°C) for at least 45 min.
constructed from the slow-learning (Ͻ30% CRs after 15 sessions)
5478 J. Neurosci., July 15, 2000, 20(14):5476–5482
Moyer et al. • Increased Excitability of Aged CA1 after Learning
Table 1. Summary of learning-related changes in CA1 neurons from young and aged rabbits
Postburst after hyperpolarization (n)
aNeurons from aged experimentally naive rabbits were significantly different from those of young naive rabbits (AHP amplitude, p Ͻ 0.005; AHP area, p Ͻ 0.01; AHP duration,
p Ͻ 0.05; number of spikes, p Ͻ 0.01). Significantly different from neurons of both age-matched control groups (naive and slow learners): *p Ͻ 0.05, ‡p Ͻ 0.01, §p Ͻ 0.001. Numbers in parentheses (n) indicate the
ratio of individual cells with reduced AHPs or accommodation to the number of cells studied in that group. Table 2. Properties of CA1 neurons from young and aged rabbits that do not change after acquisition of trace eyeblink conditioning
Action potential characteristics (n)aaAntidromic action potentials were used to allow for accurate width measurements without interference from the underlying depolarizing potential that occurs using current
injection-evoked or orthodromically driven action potentials.
and trace-conditioned rabbits clearly illustrate the poor perfor-
al., 1996b). CA1 neurons from experimentally naive aging rabbits
mance of the slow-learning rabbits from both groups (Fig. 1C, open
fired significantly fewer action potentials in response to an 800 msec
symbols) compared with rabbits that learned the task (Fig. 1C, filled
depolarizing current injection (t26 ϭ Ϫ2.988; p Ͻ 0.01) (Table 1,
symbols). Previous studies have shown that slow-learning rabbits
naive data) than did young control neurons. These data are con-
serve as an excellent control group indistinguishable from
sistent with previous observations of greater accommodation in
pseudoconditioning rabbits (Disterhoft et al., 1988b; Moyer et al.,
aging rabbit CA1 neurons compared with young adult neurons
1996). The learning curves clearly show that, although the aging
(Moyer et al., 1992; Oh et al., 1999).
rabbits were ultimately able to achieve a similar level of perfor-
No statistically significant differences were observed between
mance, throughout training they showed fewer CRs, and they took
young and aging neurons in resting membrane potential, input
nearly twice as long to reach the behavioral criterion of 60% CRs
resistance, time constant, or action potential characteristics (Table
2, naive data). In addition, the amount of current injection required
to elicit a burst of four action potentials (used to study the postburst
Aging CA1 neurons exhibit decreased postsynaptic
AHP) did not vary as a function of age (aging, 0.70 Ϯ 0.05 nA;
excitability compared with young adult CA1 neurons
Postburst AHPs of CA1 neurons from experimentally naive aged
26 ϭ 0.454; p ϭ 0.65). Analyses of within-
burst firing also indicated no statistically significant differences
rabbits were significantly larger than those from young adult neu-
between aging and young adult neurons. Latencies from current
rons. Aging CA1 pyramidal neurons had AHPs that were signifi-
onset to each of the four APs elicited during the 100 msec current
cantly larger in amplitude (t26 ϭ Ϫ3.199; p Ͻ 0.005), integrated step used to study the postburst AHP were calculated. No statisti-
area (t26 ϭ Ϫ2.871; p Ͻ 0.01), and duration (t26 ϭ 2.068; p Ͻ 0.05) cally significant differences were observed in each of the following:
than young adult CA1 neurons (Table 1, naive data). The enhanced
(1) latency to the first AP (mean, ϳ5.4 msec; t
AHPs observed in aging neurons were similar to previous reports
0.93); (2) latency to the second AP (mean, ϳ18 msec; t
of age-related changes in the AHP (Landfield and Pitler, 1984;
p ϭ 0.63); (3) latency to the third AP (mean, ϳ40 msec; t
Moyer et al., 1992). The postburst AHP is primarily comprised of
Ϫ0.901; p ϭ 0.38); or (4) latency to the fourth AP (mean, ϳ71
an outward, calcium-activated Kϩ current that modulates postsyn-
aptic excitability of many cell types, including hippocampal and
cortical pyramidal neurons (Hotson and Prince, 1980; Gustafsson
Acquisition of trace eyeblink conditioning increased
and Wigstro¨m, 1981; Lancaster and Adams, 1986; Schwindt et al.,
excitability of aging and young adult CA1 neurons
Postburst AHPs were significantly reduced in both young and aging
Spike frequency adaptation or accommodation is another mea-
CA1 neurons after acquisition of hippocampally dependent trace
sure of postsynaptic excitability (Madison and Nicoll, 1984; Hed-
eyeblink conditioning. ANOVA indicated that the effects of learn-
lund and Andersen, 1989; Moyer et al., 1992, 1996; Thompson et
ing were statistically significant for amplitude (young, F(3,46) ϭ
Moyer et al. • Increased Excitability of Aged CA1 after Learning
J. Neurosci., July 15, 2000, 20(14):5476–5482 5479 Figure 2. Acquisition of hippocampally dependent trace eyeblink conditioning increased excitability of aging rabbit hippocampal CA1 pyramidal neurons. A, Effects of trace conditioning on the size of the postburst AHP. 1, Overlay of voltage recordings of the postburst AHP in CA1 neurons from an aging
naive rabbit (Naive), an aging rabbit that showed Ͻ15% CRs after 15 sessions (Slow), and an aging trace-conditioned rabbit (Trace). The resting membrane
potentials of these cells were approximately Ϫ65 mV, with action potentials truncated for visualization of the AHP. The AHP was measured for 5 sec
beginning after a 100 msec depolarizing current injection (solid black line), with minimal current (ϳ0.6 nA) required to reliably evoke a burst of four action
potentials. 2, Mean effects of trace eyeblink conditioning on postburst AHP amplitude in aging rabbit CA1 neurons. Notice that, after learning, the AHP
was significantly reduced compared with naive and slow-learning aging controls. B, Typical examples of accommodation responses in CA1 pyramidal cells
from aging naive (Naive), aging slow-learning (Slow), and aging trace-conditioned (Trace) rabbits. Although the cell from the trace-conditioned rabbit fired
more action potentials, accommodation was not abolished, as evidenced by the increase in interspike interval with time during the 800 msec depolarizing
stimulus (solid black line), but rather was significantly reduced after learning. The resting potentials of these cells were approximately Ϫ67 mV.
21.100, p Ͻ 0.0001; aging, F(3,43) ϭ 35.382, p Ͻ 0.0001), integrated p ϭ 0.45; aging, F(3,43) ϭ 0.752, p ϭ 0.53), third (young, F(3,46) ϭ
area (young, F(3,46) ϭ 11.561, p Ͻ 0.0001; aging, F(3,43) ϭ 10.780, 0.854, p ϭ 0.47; aging, F(3,43) ϭ 0.73, p ϭ 0.54), or fourth (young,
p Ͻ 0.0001), and duration (young, F(3,46) ϭ 3.427, p Ͻ 0.05; aging, F(3,46) ϭ 1.09, p ϭ 0.36; aging, F(3,43) ϭ 1.296, p ϭ 0.3) action
F(3,43) ϭ 4.127, p Ͻ 0.01) of the AHP (Table 1). An examination of potential within each burst were not significantly different after
individual neurons indicated that, after learning, 6 of 15 (40%)
acquisition of trace eyeblink conditioning.
young adult and 17 of 19 (89%) aging neurons had significantly
After learning, CA1 neurons from both young adult and aging
reduced AHP amplitudes relative to data from age-matched naive
rabbits showed less accommodation than their age-matched control
controls (Table 1). Previous reports of learning-specific AHP re-
groups (Table 1). ANOVA revealed that significantly more action
ductions in CA1 neurons involved only the use of young adult
potentials were elicited after acquisition of trace eyeblink condi-
rabbits (Disterhoft et al., 1986; Coulter et al., 1989; de Jonge et al.,
tioning (young, F(3,46) ϭ 7.232, p Ͻ 0.001; aging, F(3,43) ϭ 10.606,
1990; Moyer et al., 1996). The present study, however, evaluated
p Ͻ 0.001). Figure 2B clearly shows this effect in aging neurons.
whether learning-related changes in postsynaptic excitability of
Notice that neurons from experimentally naive and slow-learning
CA1 neurons are also observed in aging rabbits. Figure 2 shows the
rabbits exhibited robust accommodation, whereas neurons from
effects of trace eyeblink conditioning on measures of postsynaptic
trace-conditioned rabbits fired more action potentials.
excitability of aging CA1 pyramidal neurons. The voltage traces
The learning-related changes in postsynaptic excitability (AHP
shown in Figure 2A clearly illustrate the reduced size and duration
and accommodation) were present in the absence of any statisti-
of the AHP in CA1 neurons from aging rabbits that reached the
cally significant changes in resting membrane potential, time con-
criterion of 60% CRs in a session relative to CA1 neurons from
stant, input resistance, or action potential characteristics (Table 2).
aging control rabbits (naive and slow-learning).
This was true for CA1 neurons from both young adult and aging
The learning-related changes in size and duration of the AHP
did not result from differences in current required to elicit the burst
of four action potentials used to study the postburst AHP. ANOVA
DISCUSSION
indicated that, for both young adult and aging neurons, the current
Aging rabbits were significantly slower than young adult rabbits in
required to elicit four action potentials did not vary as a function of
acquiring the trace eyeblink conditioning task. CA1 neurons from
training condition (young, F(3,46) ϭ 0.633, p ϭ 0.6; aging, F(3,43) ϭ aging naive rabbits had larger AHPs and exhibited more accom-
0.404, p ϭ 0.75). Also, there were no differences in within-
modation relative to neurons from young naive rabbits. After
burst firing in either age group as a function of training condi-
learning, both the postburst AHP and spike frequency accommo-
tion. Latencies to the first ( young, F(3,46) ϭ 0.228, p ϭ 0.88; dation were significantly reduced in a learning-specific manner in
aging, F(3,43) ϭ 1.126, p ϭ 0.35), second ( young, F(3,46) ϭ 0.903, CA1 neurons from young adult and aging rabbits. These data
5480 J. Neurosci., July 15, 2000, 20(14):5476–5482
Moyer et al. • Increased Excitability of Aged CA1 after Learning
represent the first evaluation of learning-related changes in aging
rabbits had AHP amplitudes that were 39.6% smaller after acqui-
rabbit CA1 neurons using intracellular recordings in vitro and
sition of trace eyeblink conditioning than control neurons with 40%
implicate changes in postsynaptic excitability of hippocampal neu-
of the neurons exhibiting reduced AHPs (Table 1). When given a
rons in both aging and associative learning.
long depolarizing current injection, young adult CA1 neurons fired
Aging rabbits were clearly impaired in their ability to acquire the
41.7% more action potentials after learning than did control neu-
trace eyeblink conditioning task (Fig. 1), consistent with previous
rons. Such changes were not observed in CA1 neurons from aging
observations of impaired learning ability in aging rabbits (Graves
or young rabbits that showed fewer than 30% CRs after 15 training
and Solomon, 1985; Deyo et al., 1989; Solomon and Groccia-
sessions (Table 1, Slow learners), suggesting that the effects were
Ellison, 1996; Thompson et al., 1996a). The aging rabbits required
learning-specific, as previously observed in CA1 and CA3 neurons
nearly twice as many training trials than did young adult rabbits to
from young adult rabbits (Moyer et al., 1996; Thompson et al.,
reach a behavioral criterion of 60% CRs in a session. Of the 13
aging rabbits that received trace eyeblink conditioning, only eight
Previous in vitro studies only evaluated the electrophysiological
were able to reach criterion, whereas the other five remained below
properties of aging rabbit CA1 neurons in experimentally naive
30%, even after 15 training sessions. Of the eight aging rabbits that
animals (Moyer et al., 1992; Moyer and Disterhoft, 1994; Oh et al.,
were able to learn, only two did so at a rate similar to that seen in
1999). The present data show that, although CA1 neurons from
young rabbits. These observations are consistent with previous data
aging control rabbits had larger AHPs and stronger accommoda-
indicating substantial heterogeneity of learning ability among pop-
tion than neurons from young adult controls, aging rabbits that
ulations of aging rabbits receiving trace eyeblink conditioning
learned the trace eyeblink conditioning task had AHPs that were
(Thompson et al., 1996a). In addition, the inability of the aging
significantly reduced relative to aging controls. In fact, after acqui-
rabbits probably did not result from an inability to process CS and
sition of trace conditioning, AHPs from aging rabbit CA1 neurons
US information because animals switched to the delay conditioning
were reduced to a size that was similar to that observed in the
task learn within several training sessions (Thompson et al., 1996a).
young adult rabbits after learning (Table 1). This latter point is
Hippocampal CA1 neurons recorded from experimentally naive
quite interesting because it suggests that a similar level of postsyn-
aging rabbits had significantly larger, longer lasting postburst AHPs
aptic excitability must be attained for successful acquisition of trace
(Table 1), consistent with previous reports of decreased postsyn-
conditioning, independent of the age of the animal. The actual
aptic excitability of aging rabbit and rat CA1 neurons (Landfield
differences between aging control and aging conditioned neurons
and Pitler, 1984; Moyer et al., 1992). After a burst of action
were much greater than those between young control and young
potentials, the larger, longer lasting AHPs of aging CA1 neurons
conditioned neurons (Table 1). That is, the aging neurons had to
could act to dampen the impact of excitatory inputs for several
change more than the young adult neurons to achieve the same
seconds (the duration of the AHP). Thus, a barrage of excitatory
level of postsynaptic excitability (e.g., similarly sized AHPs). The
inputs onto an aging CA1 neuron during the AHP would be less
greater change required for an aging neuron to reach the condi-
likely to drive the cell to threshold than if the cell was at or near its
tioned state may partly underlie the need for aging rabbits to
resting membrane potential. The larger AHPs observed in aging
receive significantly more training trials to successfully learn the
CA1 neurons could result from an excess influx of calcium during
trace eyeblink conditioning task. These data provide strong support
depolarization because bath application of nanomolar concentra-
for a correlation between changes in postsynaptic excitability,
tions of the L-type calcium channel antagonist nimodipine effec-
learning, and aging-related learning deficits.
tively eliminates the aging-related increase (Moyer et al., 1992).
In the present study, rabbits were trained to a behavioral crite-
Additional evidence implicating calcium or calcium-dependent
rion of 60% CRs in an 80 trial session. When rabbits were trained
processes in aging comes from work demonstrating that aging CA1
to the more difficult criterion of 80% CRs, there were little addi-
neurons have prolonged calcium action potentials (Moyer and
tional increases in CA1 postsynaptic excitability (Table 1). Al-
Disterhoft, 1994) and larger calcium currents (Landfield, 1996)
though only one aging rabbit was able to reach 80% CRs, three of
compared with young adult neurons. Preliminary data from whole-
the four cells recorded from this aging rabbit had reduced AHPs,
cell voltage-clamp experiments also suggest that the calcium-
and the mean amplitude was basically the same as those trained to
activated potassium current underlying the slow AHP is enhanced
a criteria of 60% CRs (Ϫ3.06 Ϯ 0.5 vs Ϫ3.01 Ϯ 0.2 mV, respec-
in aging rabbit CA1 neurons (Power et al., 1999).
tively). The data from young adult rabbits indicated that, when
In addition to the enhanced AHPs, CA1 neurons from aging
trained to a criterion of 80% CRs, their CA1 neurons had a mean
control rabbits also exhibited more robust accommodation during a
AHP amplitude that was only slightly smaller than those trained to
long depolarizing current injection than young adult neurons (Ta-
a criteria of 60% CRs (Ϫ2.5 Ϯ 0.3 compared with Ϫ3.02 Ϯ 0.2 mV,
ble 1) (Moyer et al., 1992). This latter observation suggests that,
respectively). Similarly, there was a slight change in accommoda-
even when aging CA1 neurons reach threshold, they are less likely
tion as a result of using a behavioral criterion of 80% CRs. On
to exhibit a sustained firing pattern in response to a continuous
average, in both age groups, training to a criterion of 80% CRs
stream of inputs. That CA1 neurons from experimentally naive
resulted in an increase of approximately one action potential dur-
aging rabbits exhibited both larger AHPs and more robust accom-
ing accommodation versus that seen when rabbits were trained to a
modation than neurons from young adult rabbits is not surprising
60% criterion (Table 1). Interestingly, there were no statistically
because modulation of the AHP by intracellular calcium or neu-
significant differences in AHP amplitude, AHP area, AHP dura-
rotransmitters typically alters accommodation (Schwartzkroin and
tion, or accommodation between young or aging CA1 neurons from
Stafstrom, 1980; Cole and Nicoll, 1983; Haas and Greene, 1984;
either trace-conditioned group. When compared with data from a
Hedlund and Andersen, 1989; Oh et al., 1999; Weiss et al., 2000).
previous study in which young adult rabbits were trained to a
After learning, CA1 neurons from aging rabbits had postburst
criterion of 80% CRs, the AHP and accommodation data observed
AHPs that were 52.5% smaller in amplitude than those from aging
24 hr after acquisition (Moyer et al., 1996) were similar to the data
control rabbits (Table 1). Inspection of individual neurons indi-
obtained in young adult CA1 neurons after conditioning to 80%
cated that, after acquisition, 89% of the aging CA1 neurons exhib-
CRs in the present study. These data suggest that the AHP reduc-
ited reduced AHPs. These effects were observed on the amplitude,
tions were nearly maximal when aged rabbits were trained to a
the integrated area, and the duration of the AHP. In response to
behavioral criterion of 60% CRs. However, in young adult rabbits,
long depolarizing current steps, CA1 neurons from aging rabbits
further reductions of the AHP occurred with additional training to
fired 76.9% more action potentials after learning than did neurons
from aging control rabbits (Table 1). These data indicate that
Additional support for involvement of changes in postsynaptic
acquisition of trace eyeblink conditioning in aging rabbits was
excitability of hippocampal CA1 neurons with learning and aging
accompanied by increased postsynaptic excitability of CA1 pyrami-
comes from studies in which compounds that reduce both the AHP
dal cells. In addition, CA1 pyramidal neurons from young adult
and accommodation were given to young adult or aging animals.
Moyer et al. • Increased Excitability of Aged CA1 after Learning
J. Neurosci., July 15, 2000, 20(14):5476–5482 5481
For example, administration of the L-type calcium channel
Haas H, Greene R (1984) Adenosine enhances AHP and accommodation
antagonist nimodipine facilitates acquisition of trace eyeblink
in hippocampal pyramidal cells. Pflu¨gers Arch 402:244–247.
conditioning in aging rabbits (Deyo et al., 1989; Straube et al.,
Hedlund B, Andersen P (1989) Alaproclate increases the excitability of
hippocampal CA1 pyramidal cells and blocks the slow after-
1990; Kowalska and Disterhoft, 1994), and nanomolar concentra-
hyperpolarization. Pharmacol Toxicol 65:185–188.
tions of nimodipine reduce both the AHP and accommodation in
Hotson JR, Prince DA (1980) A calcium-activated hyperpolarization fol-
aging rabbit CA1 neurons in vitro (Moyer et al., 1992). Similar
lows repetitive firing in hippocampal neurons. J Neurophysiol
effects have also been observed in aging rabbits treated with cho-
Khachaturian ZS (1994) Calcium hypothesis of Alzheimer’s disease and
linesterase inhibitors and muscarinic agonists (Kronforst-Collins et
brain aging. Ann NY Acad Sci 747:1–11.
al., 1997; Oh et al., 1999; Weiss et al., 2000). Compounds that
Kim JJ, Clark RE, Thompson RF (1995) Hippocampectomy impairs the
directly enhance the postburst AHP have not been tested in eye-
memory of recently, but not remotely, acquired trace eyeblink condi-
blink conditioning, but the aforementioned data suggest that such
tioned responses. Behav Neurosci 109:195–203.
Kowalska M, Disterhoft JF (1994) Relation of nimodipine dose and serum
concentration to learning enhancement in aging rabbits. Exp Neurol
Increased postsynaptic excitability appears to be one mechanism
used by hippocampal neurons in both young and aging animals for
Kronforst-Collins MA, Moriearty PL, Schmidt B, Disterhoft JF (1997)
acquisition of trace eyeblink conditioning. Previous data from
Metrifonate improves associative learning and retention in aging rabbits.
young rabbits demonstrated that changes in hippocampal excitabil-
Lancaster B, Adams PR (1986) Calcium-dependent current generating the
ity were transient, lasting 5–7 d after acquisition to a criterion of
afterhyperpolarization of hippocampal neurons. J Neurophysiol
80% CRs in a session (Moyer et al., 1996; Thompson et al., 1996b).
Although in the present study postsynaptic excitability of aging
Landfield PW (1987) “Increased calcium current” hypothesis of brain ag-
rabbit CA1 neurons was similar to young neurons after learning, it
Landfield PW (1996) Aging-related increase in hippocampal calcium
is unknown whether the increased excitability seen in aging neu-
rons would last as long as those seen in young adult neurons. The
Landfield PW, Pitler TA (1984) Prolonged Ca 2ϩ-dependent afterhyper-
current study was not designed to address this issue, but data from
polarizations in hippocampal neurons of aged rats. Science
aging rats suggest that memory consolidation (Oler and Markus,
Madison DV, Nicoll RA (1984) Control of the repetitive discharge of rat
1998) and information processing (Barnes et al., 1997; Tanila et al.,
CA1 pyramidal neurones in vitro. J Physiol (Lond) 354:319–331.
1997) are significantly impaired in aging rats.
McEchron MM, Disterhoft JF (1999) Hippocampal encoding of nonspa-
In conclusion, the present study is the first to report learning-
tial trace conditioning. Hippocampus 9:385–396.
related excitability changes in aging CA1 neurons. These data
Moyer Jr JR, Disterhoft JF (1994) Nimodipine decreases calcium action
potentials in an age- and concentration-dependent manner. Hippocam-
provide additional support for the hypothesis that alterations in
postsynaptic excitability are involved in both aging and associative
Moyer Jr JR, Deyo RA, Disterhoft JF (1990) Hippocampectomy disrupts
trace eye-blink conditioning in rabbits. Behav Neurosci 104:243–252.
Moyer Jr JR, Thompson LT, Black JP, Disterhoft JF (1992) Nimodipine
increases excitability of rabbit CA1 pyramidal neurons in an age- and
REFERENCES
concentration-dependent manner. J Neurophysiol 68:2100–2109.
Akase E, Alkon DL, Disterhoft JF (1989) Hippocampal lesions impair
Moyer Jr JR, Thompson LT, Disterhoft JF (1996) Trace eyeblink condi-
memory of short-delay conditioned eyeblink in rabbits. Behav Neurosci
tioning increases CA1 excitability in a transient and learning-specific
Akase E, Thompson LT, Disterhoft JF (1994) A system for quantitative
Norris CM, Korol DL, Foster TC (1996) Increased susceptibility to induc-
analysis of associative learning. 2. Real-time software for MS-DOS mi-
tion of long-term depression and long-term potentiation reversal during
crocomputers. J Neurosci Methods 54:119–130.
Alkon DL (1984) Calcium-mediated reduction of ionic currents: a bio-
Norris CM, Halpain S, Foster TC (1998) Reversal of age-related alter-
physical memory trace. Science 226:1037–1045.
ations in synaptic plasticity by blockade of L-type Ca 2ϩ channels. J Neu-
Barnes CA, Suster MS, Shen J, McNaughton BL (1997) Multistability of
cognitive maps in the hippocampus of old rats. Nature 388:272–275.
Oh MM, Power JM, Thompson LT, Moriearty PL, Disterhoft JF (1999)
Cole AE, Nicoll RA (1983) Acetylcholine mediates a slow synaptic po-
Metrifonate increases neuronal excitability in CA1 pyramidal neurons
tential in hippocampal pyramidal cells. Science 221:1299–1301.
from both young and aging rabbit hippocampus. J Neurosci
Coulter DA, LoTurco JJ, Kubota M, Disterhoft JF, Moore JW, Alkon DL
(1989) Classical conditioning reduces the amplitude and duration of the
Oler JA, Markus EJ (1998) Age-related deficits on the radial maze and in
calcium-dependent afterhyperpolarization in rabbit hippocampal pyra-
fear conditioning: hippocampal processing and consolidation. Hippocam-
midal cells. J Neurophysiol 61:971–981.
de Jonge MC, Black JP, Deyo RA, Disterhoft JF (1990) Learning-induced
Pitler TA, Landfield PW (1990) Aging-related prolongation of calcium
afterhyperpolarization reductions in hippocampus are specific for cell
spike duration in rat hippocampal slice neurons. Brain Res 508:1–6.
type and potassium conductance. Exp Brain Res 80:456–462.
Power JM, Oh MM, Disterhoft JF (1999) Age related enhancement of
Deyo RA, Straube KT, Disterhoft JF (1989) Nimodipine facilitates asso-
sIAHP in CA1 hippocampal pyramidal neurons. Soc Neurosci Abstr 25:84.
ciative learning in aging rabbits. Science 243:809–811.
Saar D, Grossman Y, Barkai E (1998) Reduced after-hyperpolarization in
Disterhoft JF, Kwan HH, Lo WD (1977) Nictitating membrane condition-
rat piriform cortex pyramidal neurons is associated with increased learn-
ing to tone in the immobilized albino rabbit. Brain Res 137:127–143.
ing capability during operant conditioning. Eur J Neurosci 10:1518–1523.
Disterhoft JF, Coulter DA, Alkon DL (1986) Conditioning-specific mem-
Schwartzkroin PA, Stafstrom CE (1980) Effects of EGTA on the calcium-
brane changes of rabbit hippocampal neurons measured in vitro. Proc
activated afterhyperpolarization in hippocampal CA3 pyramidal cells.
Disterhoft JF, Coulter DA, Alkon DL (1988a) Conditioning-specific bio-
Schwindt P, Spain WJ, Crill WE (1992) Calcium-dependent potassium
physical alterations in rabbit hippocampus. In: Cellular mechanisms of
currents in neurons from cat sensorimotor cortex. J Neurophysiol
conditioning and behavioral plasticity (Woody CD, Alkon DL, McGaugh
JL, eds), pp 89–104. New York: Plenum.
Shankar S, Teyler TJ, Robbins N (1998) Aging differentially alters forms of
Disterhoft JF, Golden DT, Read HL, Coulter DA, Alkon DL (1988b)
long-term potentiation in rat hippocampal area CA1. J Neurophysiol
AHP reductions in rabbit hippocampal neurons during conditioning
correlate with acquisition of the learned response. Brain Res
Solomon PR, Groccia-Ellison M (1996) Classic conditioning in aged rab-
bits: delay, trace, and long-delay conditioning. Behav Neurosci
Disterhoft JF, Moyer Jr JR, Thompson LT (1994a) The calcium rationale
in aging and Alzheimer’s disease: evidence from an animal model of
Storm JF (1990) Potassium currents in hippocampal pyramidal cells. Prog
normal aging. Ann NY Acad Sci 747:382–406.
Disterhoft JF, Moyer Jr JR, Thompson LT, Cutting FB, Power JM (1994b)
Straube KT, Deyo RA, Moyer Jr JR, Disterhoft JF (1990) Dietary nimo-
In vitro analyses of aging-related learning deficits. Soc Neurosci Abstr
dipine improves associative learning in aging rabbits. Neurobiol Aging
Graves CA, Solomon PR (1985) Age-related disruption of trace but not
Tanila H, Shapiro M, Gallagher M, Eichenbaum H (1997) Brain aging:
delay classical conditioning of the rabbit’s nictitating membrane response.
changes in the nature of information coding by the hippocampus. J Neu-
Gustafsson B, Wigstro¨m H (1981) Evidence for two types of afterhyper-
Thibault O, Landfield PW (1996) Increase in single L-type calcium chan-
polarization in CA1 pyramidal cells in the hippocampus. Brain Res
nels in hippocampal neurons during aging. Science 272:1017–1020.
Thompson LT, Deyo RA, Disterhoft JF (1990) Nimodipine enhances
5482 J. Neurosci., July 15, 2000, 20(14):5476–5482
Moyer et al. • Increased Excitability of Aged CA1 after Learning
spontaneous activity of hippocampal pyramidal cells in aging rabbits at a
excitability of rabbit CA3 neurons with a time-course appropriate to
dose that facilitates learning. Brain Res 535:119–130.
support memory consolidation. J Neurophysiol 76:1836–1849.
Thompson LT, Moskal JR, Disterhoft JF (1992) Hippocampus-dependent
Weiss C, Preston AR, Oh MM, Schwarz RD, Welty D, Disterhoft JF
learning facilitated by a monoclonal antibody or D-cycloserine. Nature
(2000) The M1 muscarinic agonist CI-1017 facilitates trace eyeblink
conditioning in aging rabbits and increases the excitability of CA1 pyra-
Thompson LT, Moyer Jr JR, Akase E, Disterhoft JF (1994) A system for
midal neurons J Neurosci 20:783–790.
quantitative analysis of associative learning. 1. Hardware interfaces with
Woody CD, Gruen E, Birt D (1991) Changes in membrane currents dur-
cross-species applications. J Neurosci Methods 54:109–117.
ing Pavlovian conditioning of single cortical neurons. Brain Res
Thompson LT, Moyer Jr JR, Disterhoft JF (1996a) Trace eyeblink condi-
tioning in rabbits demonstrates heterogeneity of learning ability both
Zyzak DR, Otto T, Eichenbaum H, Gallagher M (1995) Cognitive decline
between and within age groups. Neurobiol Aging 17:619–629.
associated with normal aging in rats: a neuropsychological approach.
Thompson LT, Moyer Jr JR, Disterhoft JF (1996b) Transient changes in
Aishima S, Nishihara Y, Iguchi T, Tagchi K, Taketomi A, Maehara Y, Tsuneypshi M. Lymphatic spread is related to VEGF-C expression and D2-40-positive myofibroblasts in intrahepatic cholangiocarcinoma. Mod Pathol 21(3):256-64,2008 Aishima S, Nishihara Y, Tsujita E, Taguchi K, Soejima Y, Taketomi A, Ikeda Y, Maehara Y, Tsuneyoshi M. Biliary neoplasia with extensive intraductal spread associated with
Kaida Y, Ueda S, Yamagishi S, Nakayama Y, Ando R, Iwatani R, Fukami K, Okuda S Proteinuria elevates asymmetric dimethylarginine levels viaprotein arginine methyltansferase-1 overexpression in a rat model of nephrotic syndrome Life Sciences Vol.91:301-305 2012 Yamagishi S, Matsui T, Ueda S, Okuda S Endocrine system Transworld Research Kajimoto H, Kai H, Aoki H, Yasuoka S, AnegawaT, Toki Y, Ue
 Moyer et al. • Increased Excitability of Aged CA1 after Learning
J. Neurosci., July 15, 2000, 20(14):5476–5482 5477
Moyer et al. • Increased Excitability of Aged CA1 after Learning
J. Neurosci., July 15, 2000, 20(14):5476–5482 5477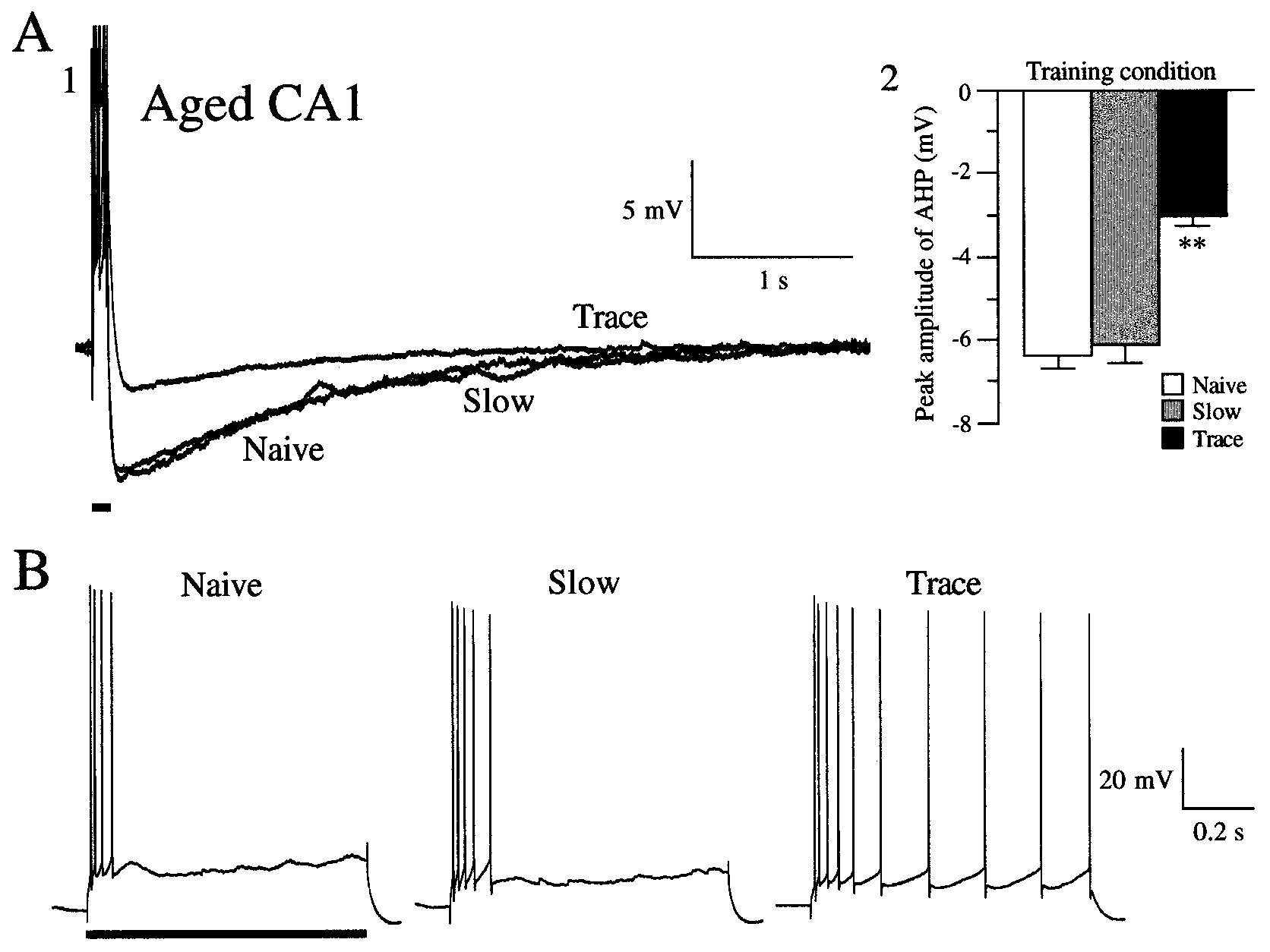 Moyer et al. • Increased Excitability of Aged CA1 after Learning
J. Neurosci., July 15, 2000, 20(14):5476–5482 5479
Moyer et al. • Increased Excitability of Aged CA1 after Learning
J. Neurosci., July 15, 2000, 20(14):5476–5482 5479