Tadalafil appartiene alla classe degli inibitori selettivi della fosfodiesterasi di tipo 5, con un profilo farmacocinetico caratterizzato da un’emivita terminale di circa diciotto ore. Dopo somministrazione orale viene assorbito rapidamente e raggiunge concentrazioni plasmatiche massime in due ore. La biotrasformazione avviene principalmente tramite CYP3A4 con formazione di metaboliti inattivi, escreti in prevalenza con le feci. L’elevato legame alle proteine plasmatiche (>90%) assicura una distribuzione stabile. Nei confronti delle altre molecole della stessa classe, cialis compresse italia è noto per la durata prolungata dell’attività farmacologica.
Microsoft word - braunmohpaperfinalversiona.doc
DUAL TUNING IN THE MAMMALIAN COCHLEA: DISSOCIATION OF NEURAL AND BASILAR MEMBRANE RESPONSES AT SUPRA- THRESHOLD SOUND LEVELS – A META-ANALYSIS Neuroscience of Music, Gansbyn 14, S-67195 Klässbol, Sweden
In the basal turn of the cochlea, tuning of auditory nerve fiber (ANF) and basilar membrane (BM) responses are closely correlated at threshold levels. Their correlation also at supra-threshold levels has always been an implicit postulate of BM transmission line theory. A test of this postulate by empirical data, however, has never been published. In order to elucidate this discrepancy, a large number of ANF recordings was analyzed and compared to equivalent BM recordings. The results show that, at least in the basal turn of the cochlea, the best frequency (BF) of BM responses drops by about half an octave between 30 and 100 dB SPL. The comparable neural data never reflect any-thing similar. 1) In the majority of cases (52%) BF is totally unaffected by sound level. 2) In a mi-nority of cases (36%) BF drops slightly at the highest sound levels, but only by about half as much as for the BM. 3) In a relevant number of cases BF is bimodal at the highest sound levels, such that the antagonistic majority and minority versions appear concurrently. These results indicate a dual tuning, such that BM and organ of Corti (OC) are tuned separately, and they are consistent with the concept of overload protection of OC sensitivity as the genuine function of BM tuning.
Introduction
For ten years now, we know that in the mammalian cochlea responses of the basilar mem-brane (BM) and of auditory nerve fibers (ANFs) can be closely related at threshold of hearing [1, 2]. As usual, however, correlation says nothing about cause and effect. Cur-rently, there are two competing and mutually exclusive schools of thought. The first one suggests that at threshold of hearing the chain of activation is BM-OHC-BM-IHC-ANF [3]. The second one suggests that it is OHC-IHC-ANF [4, 5], with the simultaneous OHC-BM branch as an epiphenomenon [6, 7]. Important information on the matter can be ex-pected from data on BM and ANF responses at supra-threshold sound levels.
If these are correlated like they are at threshold (outcome A), the second hypothesis
(“OHC first”) will be unrealistic, because at high sound levels the relative contribution of OHC motility to mechanical forces in the cochlea is too weak to dominate BM behavior.
If BM and ANF responses are not correlated at supra-threshold sound levels (out-
come B), the first hypothesis (“BM first”) will be unrealistic, because indications for a chain of activation BM-IHC-ANF will then be missing.
From the published literature two bodies of evidence were scrutinized: (a) all data on level dependence of the best frequency (BF) of single ANFs that appeared after the intro-duction of single fiber recording in this field in the mid-1960s, and (b) all data on level dependence of BF of BM responses that appeared after the introduction of laser interfero-
metry in this field in 1990. The major trends within each of the two data bodies were de-termined and then compared across data bodies.
3.1. Basilar membrane
For the basal turn of non-specialized mammalian cochleae sufficient data are available, and the results are uniform. A typical example is shown in Fig. 1. Here BF is stable at 9.5 kHz from 0 to 20 dB, but already at 30 dB it has migrated to 9.25 kHz. The migration toward lower BFs then continues monotonously, reaching 9.0 kHz at 50 dB, 8.5 kHz at 70 dB, 7.5 at 80 dB, and 6.5 at 100 dB. In octaves (oct) the cumulated shifts are 0.04, 0.08, 0.16, 0.34, and 0.55, respectively.
3.2. Auditory nerve fibers
There are two large-scale investigations of the sound-level dependence of ANF firing rates in the squirrel monkey, by Rose et al. [8] (R71) and by Geisler et al. [9] (G74). Both studies can be considered as normative, because of (a) the very large quantity of the col-lected and published data and (b) the absence of deviating results elsewhere in the litera-ture.
R71 recorded data from 48 animals and published 14 figures with iso-intensity
curves. In 13 of them BF did not vary with sound level. In one (Fig. 2B) BF shifted slightly toward lower frequencies, from 2.1 to 1.7 kHz (0.30 oct) in the range from 50 to 90 dB.
G74 recorded data from 31 animals and published 9 figures with iso-intensity curves.
In six of them BF did not vary with sound level. In two (Figs. 2A , 2B) BF shifted slightly toward lower frequencies, from 4.6 to 4.3 kHz (0.10 oct) in the range from 50 to 80 dB and from 7.3 to 6.9 kHz (0.08 oct) in the range from 50 to 80 dB. In one (Fig. 8) BF shifted slightly toward higher frequencies, from 3.5 to 3.7 kHz (0.08 oct) in the range from 65 to 95 dB. Further, G74 stated the proportional distribution of types of BF shifts as present in their total data base: “52 % of the fibers sampled showed no change in best frequency with intensity changes” (p. 1161); in 36 % BF dropped “slightly”, and in 12 % it increased with level.
In five of the nine iso-intensity plots of G74 BF behavior is clearly visible even at the
highest sound levels, i.e. not hidden by high-level saturation. In four of these five, BF shows a bimodal behavior at highest levels. Three examples are displayed in Fig. 1, where the broken lines in the ANF plots indicate secondary BFs on the low-frequency side, with deviations from the normal BFs of 0.21, 0.21, and 0.26 oct, respectively, at 100 dB.
3.3. Comparison BM versus ANF
1) The majority of ANFs has a stable BF across sound levels and shows no relation at all to the BM’s shift of BF.
Figure 1. BF versus sound level. X-axis: BF as defined by the peaks in the iso-intensity curves of BM and ANF responses. Y-axis: sound intensity of the stimulus. The four curves are spaced in half-octaves re threshold BF, which is stated in kHz below each curve. Leftmost curve: plotted from the BM velocity data in Fig. 1A of Ruggero et al. [2]. Other three curves: plotted from the ANF firing-rate data (3 fibers of 2 animals) in Figs. 7A-C of Geisler et al. [9]. Stimuli were 128 to 512 tone bursts of 5 to 100 ms per stimulus condition for the BM recordings and 4 to 8 tones of 500 ms per stimulus condition for the ANF recordings.
Figure 2. Cochlear frequency mapping versus sound level. X-axis: BF place position along the length of the cochlea. Y-axis: sound intensity of the stimulus. BM data according to sample in Fig. 1. ANF data according to the summarized results of the present study.
2) A minority of ANFs shows a BF shift at the highest sound levels that goes into the same direction as observed at the BM, but is only about half as large. 3) Some ANFs show a bimodal BF at the highest sound levels, with one mode at the fi-ber’s normal BF and the other mode reflecting a shift that goes into the same direction as observed at the BM, but is only about half as large.
3.4. Stability of BF of ANF responses in cross-correlation data
Bimodal BFs, similar to those in Fig. 1, have also been documented in cross-correlation analyses of ANF responses to broadband noise at high sound levels [10; Fig. 3]. Further, since 2005 we have data from Wiener-kernel analysis of ANF responses to broadband noise even from the basal cochlear turn [11; Fig. 14]. Because these data are from the same species and from a similar frequency place as the BM data in Fig. 1, a direct com-parison is possible. For sound levels from 33 to 68 dB SPL in ERB the BF shift of the ANF is very small: from 12.1 to 11.8 kHz (0.036 oct). The comparable BF shift of the BM (Fig. 1), however, is more than three times as large: from 9.25 to 8.5 kHz (0.12 oct).
3.5. Level independence of BF in further neural pathway
Concerning the further neural pathway, a BF shift that is similar to the one in BM re-sponses has never been reported. To the contrary, in the cochlear nucleus it is common that neurons which apparently receive their major excitatory input from the auditory nerve have a strictly constant BF, even if their dynamic range is >80 dB [12; Fig. 3C]. Also for the inferior colliculus the level independence of BF of sharply tuned neurons is well docu-mented [13; Figs. 2 & 11].
Discussion 4.1. Dual tuning
The finding of a clear dissociation between BM tuning and ANF tuning at medium and high sound levels indicates a dual mechanism. Besides the well-known BM tuning, which also occurs post mortem at levels above ca 60 dB, there appears to be an independent tuning mechanism in the organ of Corti (OC) that underlies ANF behavior.
The fact that a minority of ANFs partially, but never fully, follows the BF shift of the
BM further confirms a dual mechanism. It can be understood as an effect of the well-known phenomenon of frequency pulling in the cochlea [14]. Limit cycle oscillators in the OC, as a possible cause of the BF of ANFs, could under certain conditions be partially entrained by strong nearby BM oscillations.
4.2. Function of dual tuning
The dissociation between BM and ANF tuning at supra-threshold sound level is also
reflected in the well-documented phenomenon of the “half-octave shift in temporary threshold shift (TTS)” [15]. Here, exposure to a high-level tone typically reduces hearing
sensitivity at a frequency half an octave above the exposure frequency, while leaving sensitivity fully intact at the exposure frequency itself. This dissociation of exposure fre-quency and frequency range of damage precisely mirrors the dissociation of BM and neural tuning. Most importantly, the “half-octave shift in TTS” protects sensitivity at the exposure frequency. As shown in Fig. 2, a section of the BM that lies basalward of a given characteristic frequency (CF) absorbs vibrational energy before this energy reaches the place of CF. Thus, the present results further substantiate earlier observations that overload protection appears to be the genuine function of the BM traveling wave in the mammalian inner ear [4, 16, 17].
Conclusions
1) The “BM-first” hypothesis of cochlear activation is not compatible with the present findings, because BM tuning dissociates from OC tuning at sound levels above 20 dB SPL. 2) The “OHC-first” hypothesis of cochlear activation is compatible with the present find-ings, because in the sound level range that is dominated by OHC motility the apparent BM tuning is almost congruent with OC tuning. 3) The BM’s passive tuning at medium and high sound levels is strategically placed for a protection of the sensitivity of the OC’s active tuning, because it absorbs energy basal-ward of any given CF in the OC.
Addendum
After the completion of this study, Mellado Lagarde et al. [18] reported data on the disso-ciation between BM and neural responses that are in remarkable agreement with the pre-sent results and conclusions. In mice where the gene for the OHC motor protein prestin was deleted CF of BM responses was reduced by half an octave. Further, neural responses were less sensitive by >50 dB SPL than BM responses. These new data appear to reflect the same co-occurrence of two separate tuned mechanisms as described in the present study, because both the half-octave disparity and a striking independence of ANF and BM responses were observed again. Also here indications for a chain of activation BM-IHC-ANF were missing, and thus the “BM-first” hypothesis again appeared to be unrealistic.
References
1. Narayan, S.S., Temchin, A.N., Recio, A., Ruggero, M.A., 1998. Frequency tuning of
basilar membrane and auditory nerve fibers in the same cochleae. Science 282, 1882-1884.
2. Ruggero, M.A., Narayan, S.S., Temchin, A.N., Recio, A., 2000. Mechanical bases of
frequency tuning and neural excitation at the base of the cochlea: comparison of basi-lar-membrane vibrations and auditory-nerve fiber responses in chinchilla. Proc. Natl. Acad. Sci. USA 97, 11744-11750.
3. Robles, L., Ruggero, M.A., 2001. Mechanics of the mammalian cochlea. Physiol.
4. Braun, M., 1994. Tuned hair cells for hearing, but tuned basilar membrane for over-
load protection: evidence from dolphins, bats, and desert rodents. Hear. Res. 78, 98-114.
5. Nowotny, M., Gummer, A.W., 2006. Nanomechanics of the subtectorial space
caused by electromechanics of cochlear outer hair cells. Proc. Natl. Acad. Sci. USA 103, 2120-2125.
6. Dancer, A., Avan, P., Magnan, P., 1997. Can the traveling wave be challenged by
direct intra-cochlear pressure measurements? In: Lewis, E.R., Long, G.R., Lyon, R.F., Narins, P.M., Steele, C.R., Hecht-Poinar, E. (Eds.), Diversity in Auditory Me-chanics. World Scientific, Singapore, pp. 340-346.
7. Pujol, R., Eybalin, M., Lavigne-Rebillard, M., Lenoir, M., Puel, J.L., Rebillard, G.,
Ruel, J., Trigueiros-Cunha, N., Wang, J., 1999-2007. Promenade around the Coch-lea. Section “Cochlea”, subsection “physics”. http://www.cochlea.org/
8. Rose, J.E., Hind, J.E., Anderson, D.J., Brugge, J.F., 1971. Some effects of stimulus
intensity on response of auditory nerve fibers in the squirrel monkey. J. Neuro-physiol. 34, 685-699.
9. Geisler, C.D., Rhode, W.S., Kennedy, D.T., 1974. Responses to tonal stimuli of sin-
gle auditory nerve fibers and their relationship to basilar membrane motion in the squirrel monkey. J. Neurophysiol. 37, 1156-1172.
10. Møller, A.R., 1978. Frequency selectivity of the peripheral auditory analyzer studied
using broad band noise. Acta. Physiol. Scand. 104, 24-32.
11. Recio-Spinoso, A., Temchin, A.N., van Dijk, P., Fan, Y.H., Ruggero, M.A., 2005.
Wiener-kernel analysis of responses to noise of chinchilla auditory-nerve fibers. J. Neurophysiol. 93, 3615-3634.
12. Spirou, G.A., Davis, K.A., Nelken, I., Young, E.D., 1999. Spectral integration by
type II interneurons in dorsal cochlear nucleus. J. Neurophysiol. 82, 648-663.
13. Ramachandran, R., Davis, K.A., May, B.J., 1999. Single-unit responses in the infe-
rior colliculus of decerebrate cats. I. Classification based on frequency response maps. J. Neurophysiol. 82, 152-163.
14. Long, G.R., Tubis, A., Jones, K.L., 1991. Modeling synchronization and suppression
of spontaneous otoacoustic emissions using Van der Pol oscillators: Effects of aspirin administration. J. Acoust. Soc. Am. 89, 1201-1212.
15. McFadden, D., 1986. The curious half-octave shift: evidence for a basalward migra-
tion of the traveling-wave envelope with increasing intensity. In: Salvi, R.J., Hender-son, D., Hamernik, R.P., Coletti, V. (Eds.), Basic and Applied Aspects of Noise-induced Hearing Loss. Plenum Publishing, New York, pp. 295-312.
16. Braun, M., 1993. Basilar membrane tuning re-examined: frequency selective damp-
ing of high level input may be its genuine function. In: Duifhuis, H., Horst, J.W., van Dijk, P., and van Netten, S.M. (Eds.), Biophysics of Hair Cell Sensory Systems. World Scientific, Singapore, p. 406.
17. Braun, M., 1996. Impediment of basilar membrane motion reduces overload protec-
tion but not threshold sensitivity: evidence from clinical and experimental hydrops. Hear. Res. 97, 1-10.
18. Mellado Lagarde, M.M., Drexl, M., Lukashkin, A.N., Zuo, J., Russell, I.J., 2008.
Prestin's role in cochlear frequency tuning and transmission of mechanical responses to neural excitation. Curr. Biol. 18, 200-202.
BIJSLUITER: INFORMATIE VOOR DE GEBRUIKER DEXTROMETHORFAN QUALIPHAR, 1 mg/ml, siroop Dextromethorfan hydrobromide Lees goed de hele bijsluiter voordat u dit geneesmiddel gaat innemen want er staat belangrijke informatie in voor u. Gebruik Dextromethorfan Qualiphar altijd precies zoals beschreven in deze bijsluiter of zoals uw arts of Dit geneesmiddel kunt u zonder voorschrift kr
Intended use The ßhCG test is indicated for use as an aid in the early detection of pregnancy. The test is not indicated as a surrogate marker in the diagnosis or monitoring of cancer patients. SummaryHuman chorionic gonadotropin (hCG) is a glycoprotein hormone. It is secreted during pregnancy by the trophoblastic cells of the placenta, shortly after the implantation of the fertilized ovum in
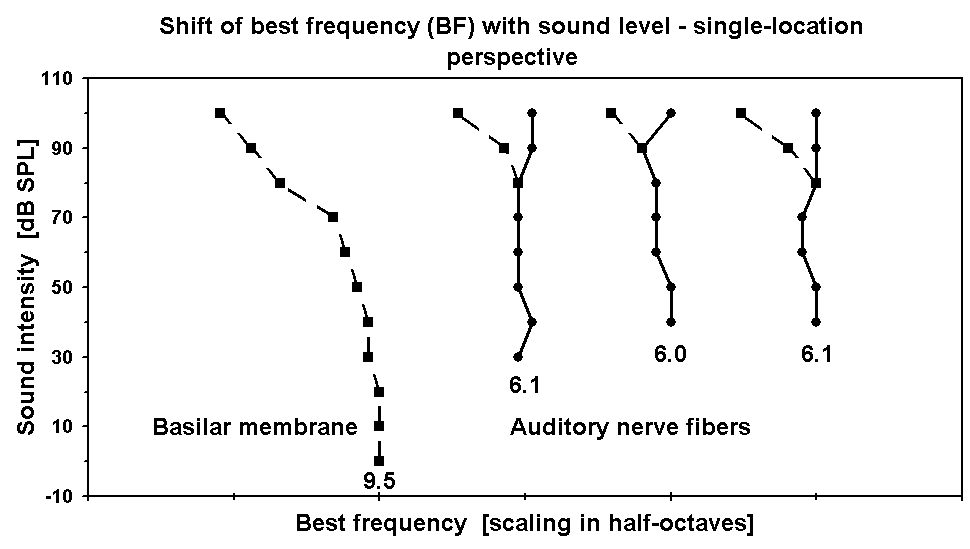
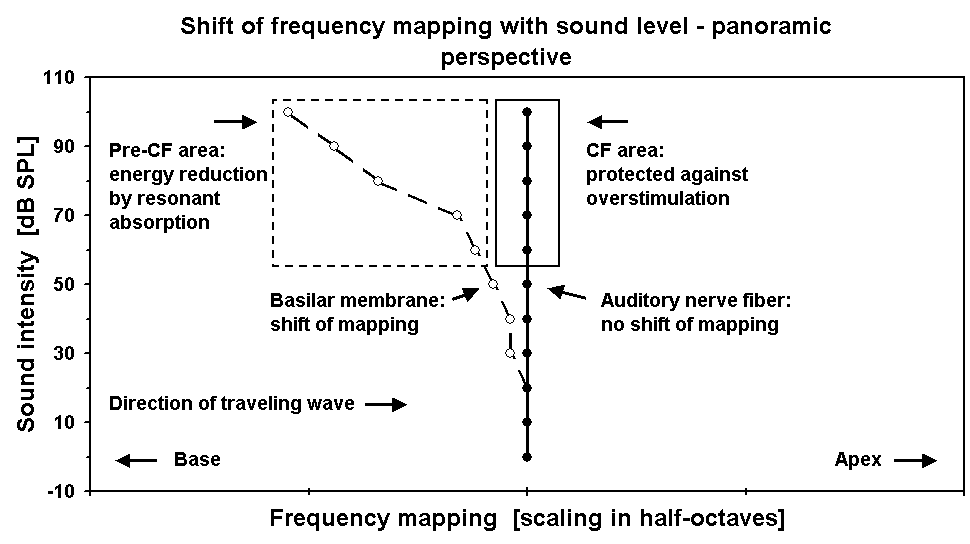 Figure 1. BF versus sound level. X-axis: BF as defined by the peaks in the iso-intensity curves of BM and ANF responses. Y-axis: sound intensity of the stimulus. The four curves are spaced in half-octaves re threshold BF, which is stated in kHz below each curve. Leftmost curve: plotted from the BM velocity data in Fig. 1A of Ruggero et al. [2]. Other three curves: plotted from the ANF firing-rate data (3 fibers of 2 animals) in Figs. 7A-C of Geisler et al. [9]. Stimuli were 128 to 512 tone bursts of 5 to 100 ms per stimulus condition for the BM recordings and 4 to 8 tones of 500 ms per stimulus condition for the ANF recordings.
Figure 2. Cochlear frequency mapping versus sound level. X-axis: BF place position along the length of the cochlea. Y-axis: sound intensity of the stimulus. BM data according to sample in Fig. 1. ANF data according to the summarized results of the present study.
2) A minority of ANFs shows a BF shift at the highest sound levels that goes into the same direction as observed at the BM, but is only about half as large. 3) Some ANFs show a bimodal BF at the highest sound levels, with one mode at the fi-ber’s normal BF and the other mode reflecting a shift that goes into the same direction as observed at the BM, but is only about half as large.
3.4. Stability of BF of ANF responses in cross-correlation data
Figure 1. BF versus sound level. X-axis: BF as defined by the peaks in the iso-intensity curves of BM and ANF responses. Y-axis: sound intensity of the stimulus. The four curves are spaced in half-octaves re threshold BF, which is stated in kHz below each curve. Leftmost curve: plotted from the BM velocity data in Fig. 1A of Ruggero et al. [2]. Other three curves: plotted from the ANF firing-rate data (3 fibers of 2 animals) in Figs. 7A-C of Geisler et al. [9]. Stimuli were 128 to 512 tone bursts of 5 to 100 ms per stimulus condition for the BM recordings and 4 to 8 tones of 500 ms per stimulus condition for the ANF recordings.
Figure 2. Cochlear frequency mapping versus sound level. X-axis: BF place position along the length of the cochlea. Y-axis: sound intensity of the stimulus. BM data according to sample in Fig. 1. ANF data according to the summarized results of the present study.
2) A minority of ANFs shows a BF shift at the highest sound levels that goes into the same direction as observed at the BM, but is only about half as large. 3) Some ANFs show a bimodal BF at the highest sound levels, with one mode at the fi-ber’s normal BF and the other mode reflecting a shift that goes into the same direction as observed at the BM, but is only about half as large.
3.4. Stability of BF of ANF responses in cross-correlation data